Click image to enlarge with full caption. Main text below slider.

01 Tritia reticulata

02 Tritia reticulata

03 Tritia reticulata

04 Tritia reticulata

05 Tritia reticulata

06 Tritia reticulata

07 Tritia reticulata

08 Tritia reticulata
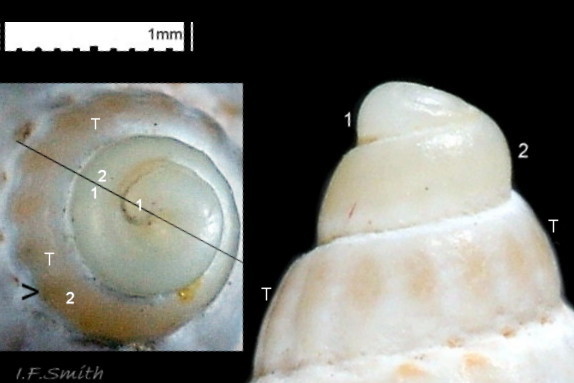
09 Tritia reticulata

10 Tritia reticulata

11 Tritia reticulata
12 Tritia reticulata

13 Tritia reticulata

14 Tritia reticulata

15 Tritia reticulata
16 Tritia reticulata

17 Tritia reticulata

18 Tritia reticulata

19 Tritia reticulata

20 Tritia reticulata
21 Tritia reticulata
22 Tritia reticulata

23 Tritia reticulata
24 Tritia reticulata
25 Tritia reticulata
26 Tritia reticulata
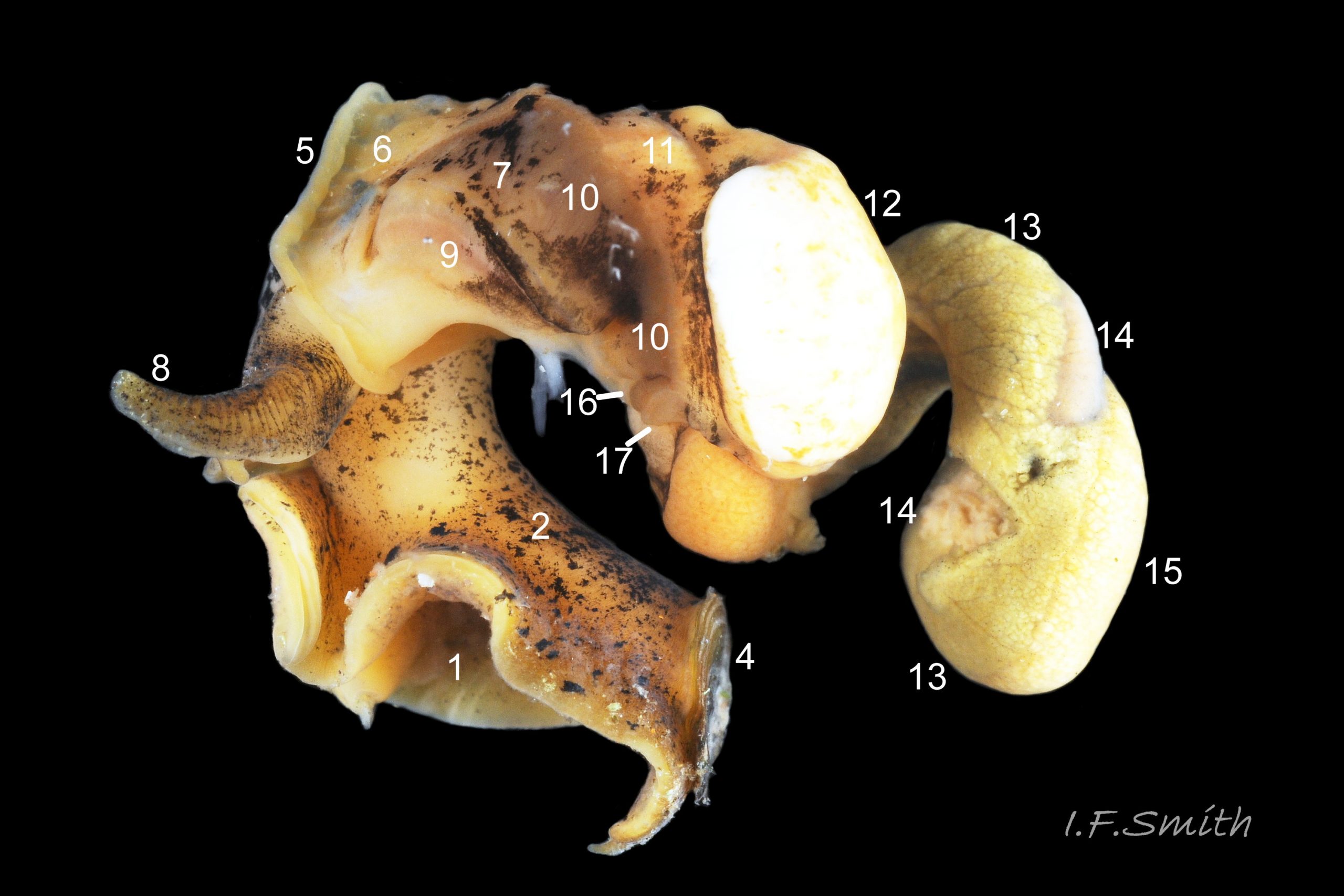
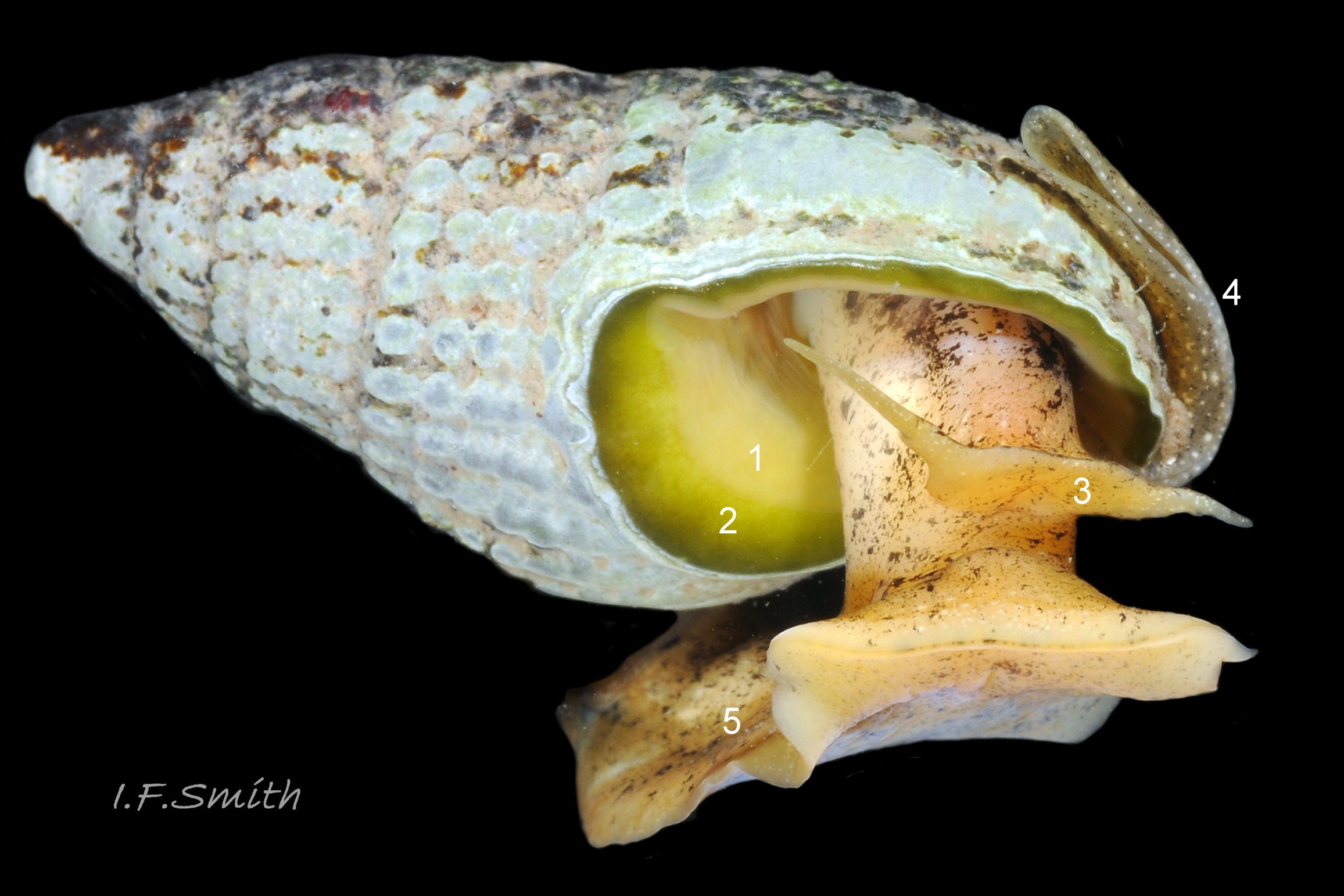
27 Tritia reticulata. Dissected female, mantle intact. Llŷn Penisula, Wales. March 2016.

28 Tritia reticulata

29 Tritia reticulata

30 Tritia reticulata
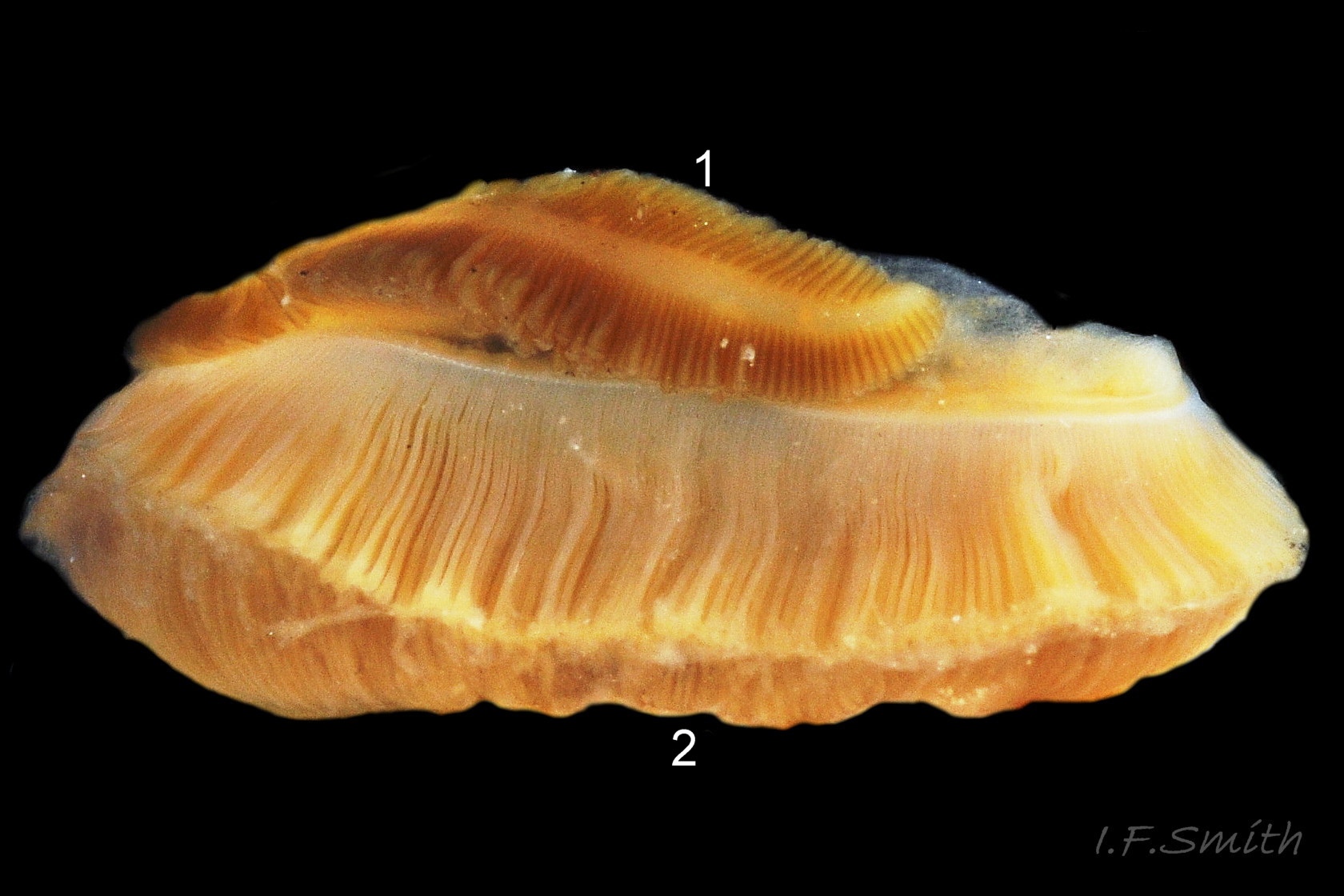
31 Tritia reticulata

32 Tritia reticulata

33 Tritia reticulata

34 Tritia reticulata

35 Tritia reticulata
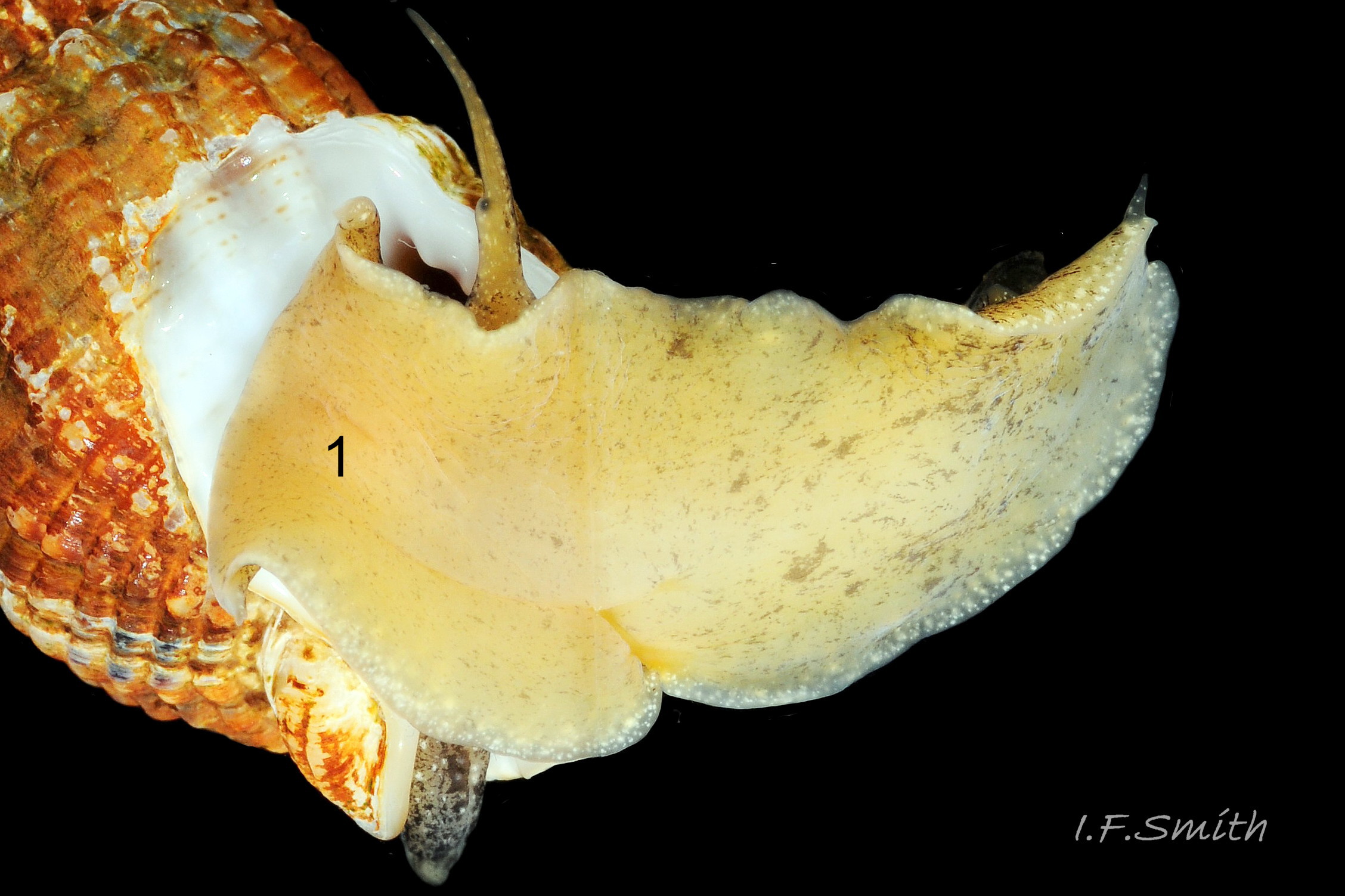
36 Tritia reticulata

37 Tritia reticulata

38 Tritia reticulata

39 Tritia reticulata COMPARE Tritia nitida

40 Tritia reticulata

41 Tritia reticulata

42 Tritia reticulata
Predatory Euspira spp. may bore the shells of young T. reticulata near the apex where thinnest, but below the persistent protoconch.
Shell height 10 mm. Llŷn, North Wales, August, 1968.

43 Tritia reticulata

44 Tritia reticulata

45 Tritia reticulata

46 Tritia reticulata COMPARE Tritia nitida,
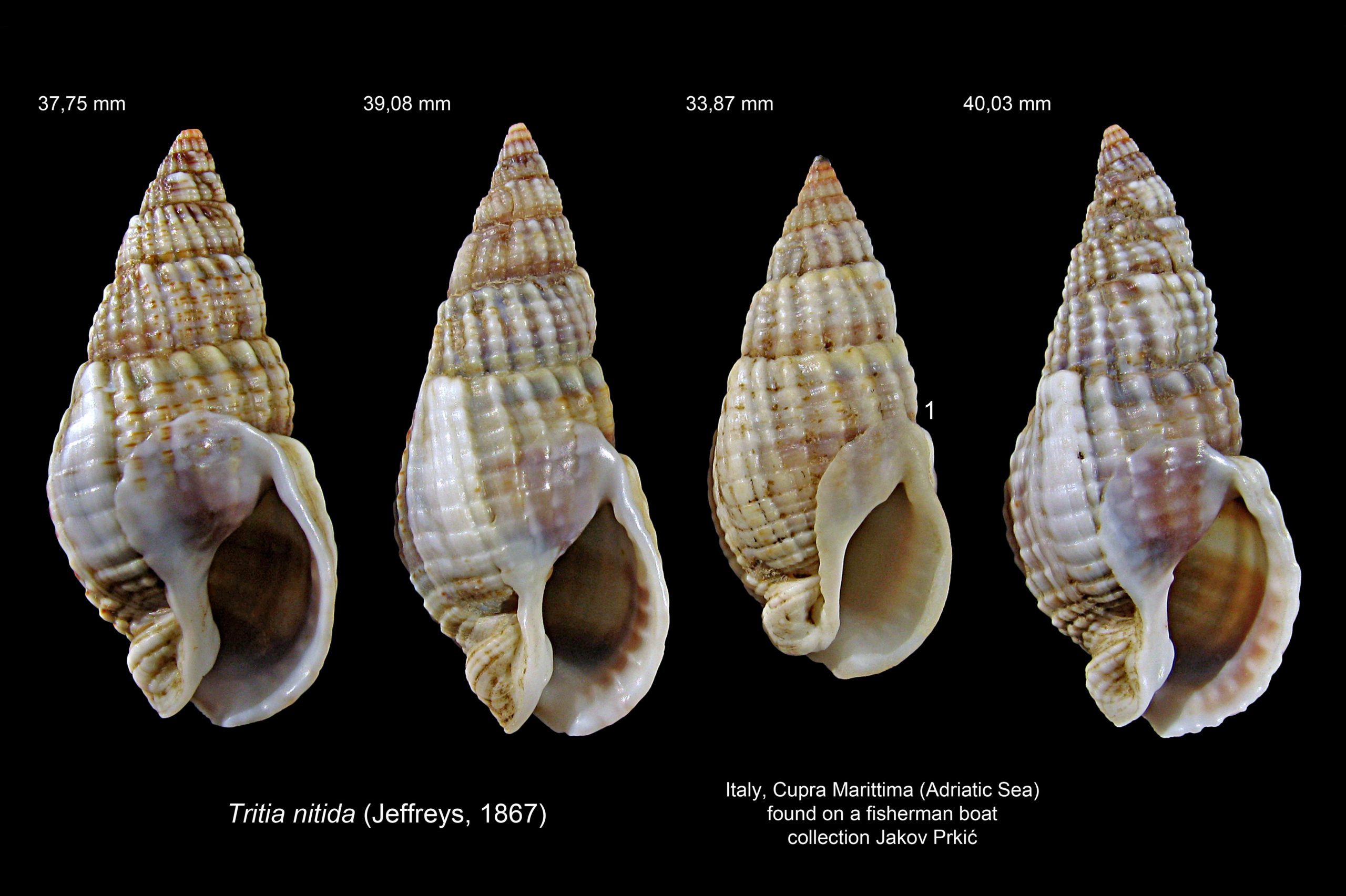
47 Tritia reticulata COMPARE Tritia nitida

48 Tritia reticulata COMPARE Tritia nitida

49 Tritia reticulata
Concentration of T. reticulata individuals attracted by the scent of rotting to feed on carrion. Aughrus, Connemara, Ireland. June, 2017. © M. Thomas.

50 Tritia reticulata COMPARE Tritia nitida

51 Tritia reticulata COMPARE Tritia nitida

52 Tritia reticulata COMPARE Tritia nitida

53 Tritia reticulata COMPARE Tritia nitida

54 Tritia reticulata COMPARE Tritia nitida

55 Tritia reticulata COMPARE Tritia nitida

56 Tritia reticulata COMPARE Tritia nitida
In Britain the body whorl has 15 to 23 costae on T. reticulata and 10 to 14 on T. nitida, but in the Adriatic the range on T. nitida is wider, 9 to 18, and overlaps with that of T. reticulata.
<!– wp:paragraph …
57 Tritia reticulata COMPARE Tritia nitida

58 Tritia reticulata COMPARE Tritia nitida

59 Tritia reticulata COMPARE Tritia nitida
60 Tritia reticulata COMPARE Tritia nitida

61 Tritia reticulata COMPARE Tritia nitida

62 Tritia reticulata COMPARE Tritia nitida

63 Tritia reticulata COMPARE Tritia nitida

64 Tritia reticulata COMPARE Tritia nitida

65 Tritia reticulata COMPARE Tritia nitida
- nitida egg capsules are slenderly ovoid with W/H 65% including neck when recently laid, mean height 2.65 mm, width 1.72 mm (Roland & Luque, 1994).

66 Tritia reticulata COMPARE Tritia incrassata

67 Tritia reticulata COMPARE Tritia incrassata

68 Tritia reticulata COMPARE Tritia incrassata

69 Tritia reticulata COMPARE Tritia varicosa

70 Tritia reticulata COMPARE Tritia varicosa
Tritia reticulata (Linnaeus, 1758)
Synonyms: Buccinum reticulatum Linnaeus, 1758; Hinia reticulata (Linnaeus, 1758); Nassarius reticulatus (Linnaeus, 1758); Nassa reticulata (Linnaeus, 1758).
Current taxonomy: World Register of Marine Species (WoRMS) https://www.marinespecies.org/aphia.php?p=taxdetails&id=876821
Vernacular: Netted dog whelk (English); Chwalcen rwyllog (Welsh); Gevlochten fuikhoren (Dutch); Netzreusenschnecke (German); Nasse réticulée (French); Dværgkonk (Danish); Κοφίνι (Greek).
GLOSSARY below.
Shell description.
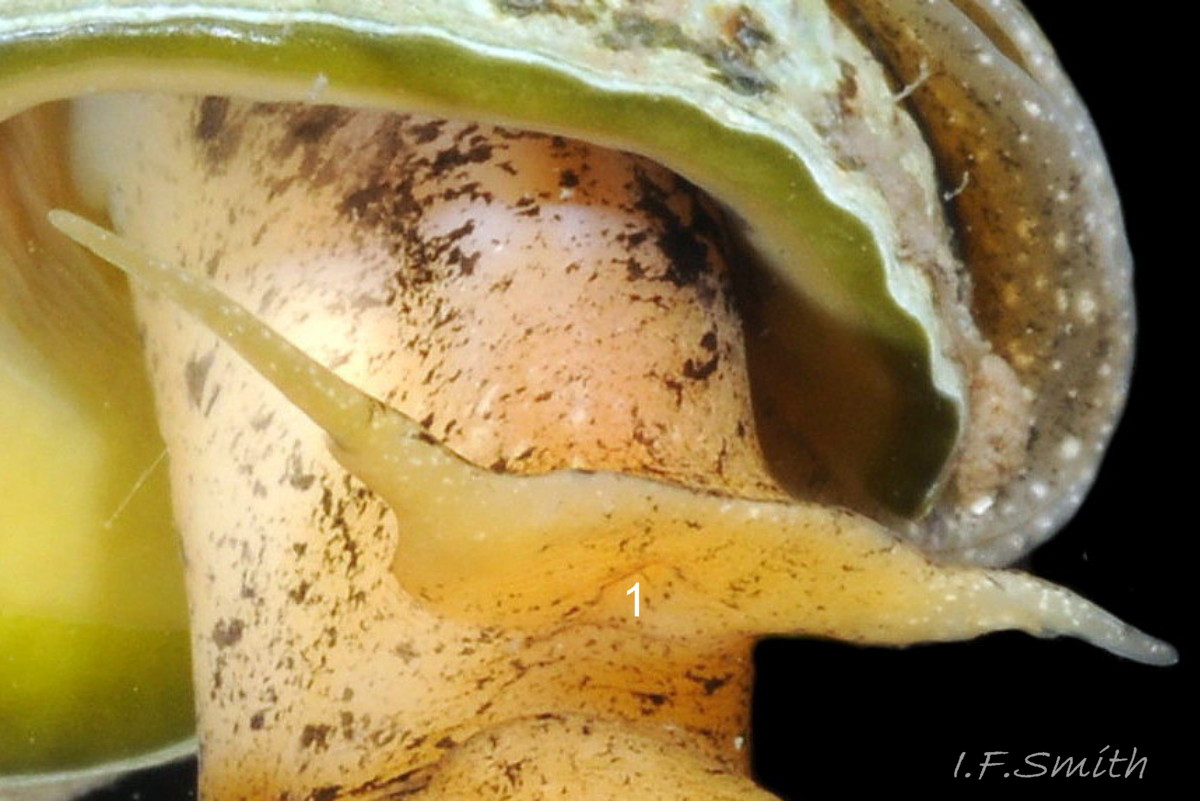
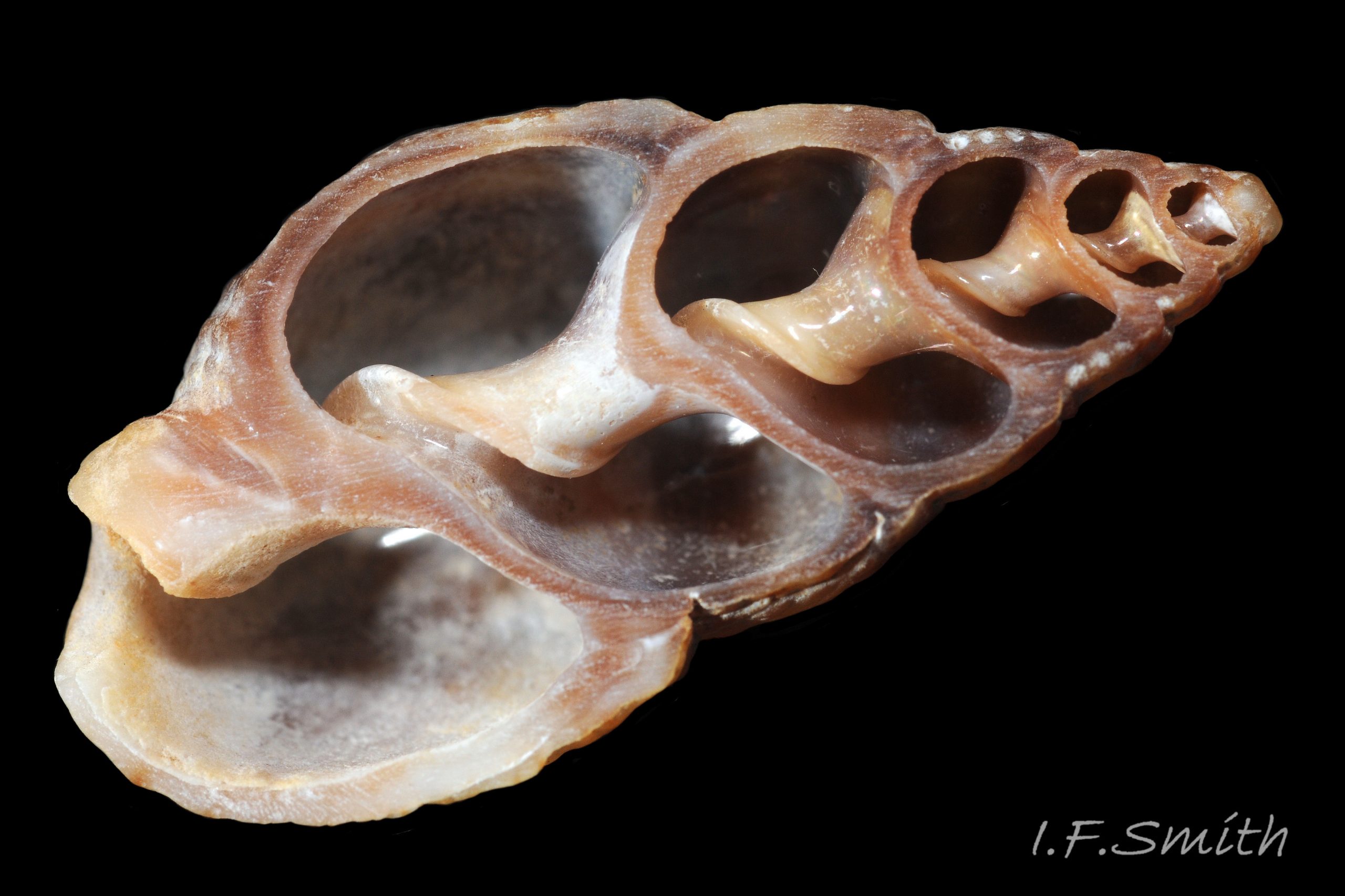
The shell of T. reticulata grows to a maximum of 30 mm high and 14 mm wide. The shell wall is solid, opaque and matt. The body whorl is convex but the spire whorls are almost flat and the spire profile is almost straight sided 01 Tritia reticulata . Juveniles may have a slightly concave spire profile 02 Tritia reticulata . The sutures are shallow, which is sometimes emphasised by each whorl being raised slightly higher than the preceding whorl 01 Tritia reticulata. The spire is a variably slender, sharply pointed cone with adult width varying from 45% to 55% of height 01 Tritia reticulata . Juvenile spires taper more sharply with width/height about 55%, or more 02 Tritia reticulata . Adult body whorl height is 60% to 70% of shell height 03 Tritia reticulata . The shell has a sculpture of broad spiral ridges, narrower spiral grooves, and sigmoid costae about as width as the intervening gaps. In Britain, the adult body whorl has 15 to 23 costae (pers. obs. IFS, 2019) or 15 to 20 (Jeffreys, 1867). In Ria de Vigo, north-west Spain, 16 to 23 costae were recorded by Rolan and Luque (1994). Juveniles may have more costae. Rarely, the final costa is sufficiently thickened to form a slight labial varix 04 Tritia reticulata . The exterior rim of the aperture is usually thin, smooth and white without periostracum 05 Tritia reticulata & 06 Tritia reticulata . The intersection of costae and spiral ridges creates rectangular bosses in a reticulate arrangement 07 Tritia reticulata . There is a deep spiral gutter at the base of the shell. Below the gutter, the neck of the siphonal canal has 4 or 5 grooves on its exterior when not eroded 07 Tritia reticulata . The protoconch has a diameter of about 1 mm and is smooth and white 08 Tritia reticulata with a little more than 2 whorls (Sanjuan, 1997). In Ria de Vigo, Sanjuan (1997) found this to differentiate it from T. nitida with a little less than two complete whorls, but some in Britain seem to have 1.8 whorls on the protoconch 09 Tritia reticulata . In the Ria de Vigo, Sanjuan (1997) found the protoconch to persist on most live T. reticulata and to be incomplete on most T. nitida. This applied generally also to British material examined by IFS, the apical whorls being frequently broken off on T. nitida and often persisting to form a fine point, even when worn, on T. reticulata 10 Tritia reticulata. When live and uneroded, the exterior of the shell is covered by a brown periostracum apart from the protoconch and the rim of the outer (palatal) lip 05 Tritia reticulata . On worn shells the periostracum persists longest in the shelter of the grooves between the costae and spiral ridges 11 Tritia reticulata .
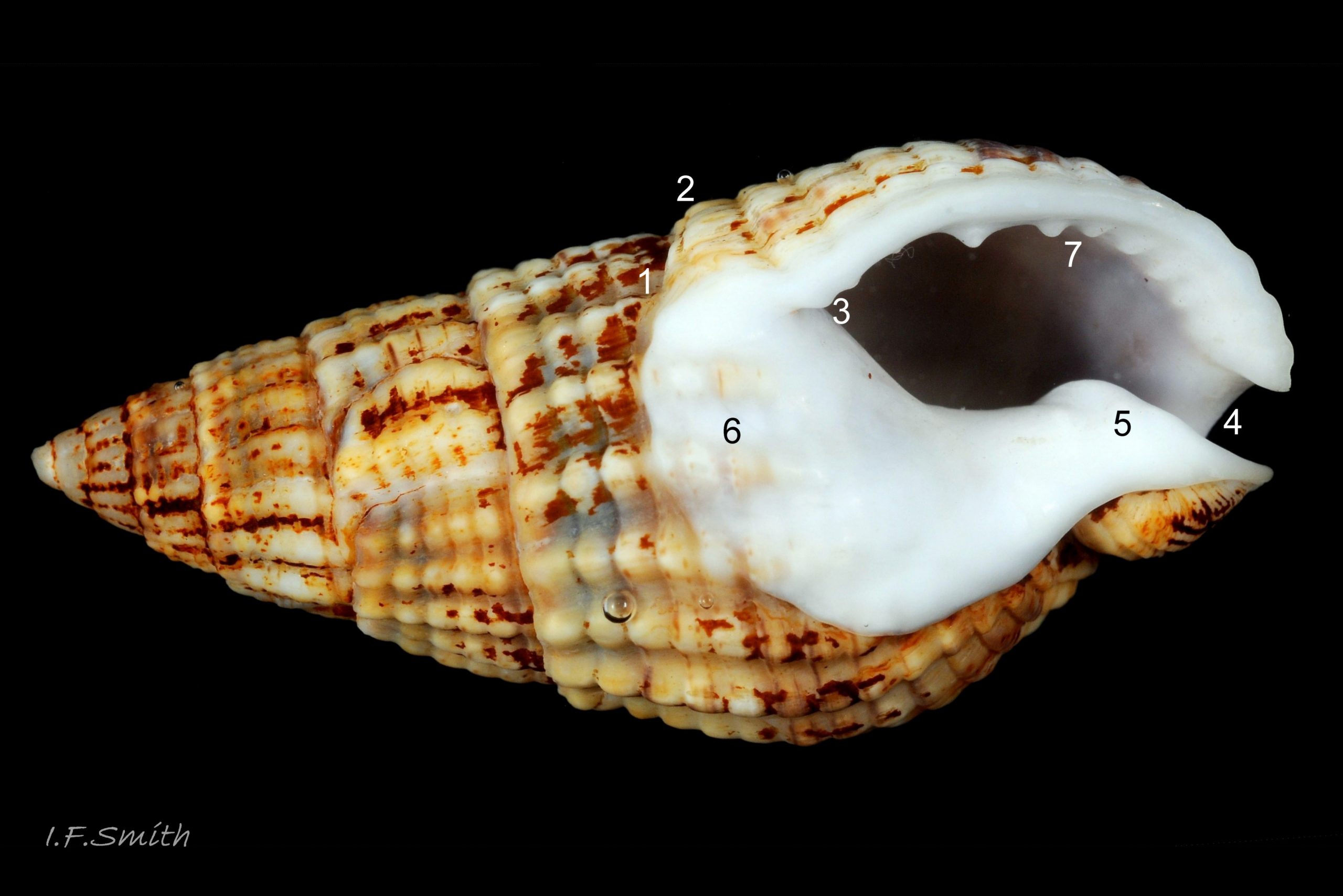
The height of the oval aperture is about 45% of the shell height 03 Tritia reticulata . The palatal (outer) lip meets the parietal (upper, inner) lip at about 90º (not always discernible) but immediately curves strongly abapically so the aperture is pointed adapically12 Tritia reticulata . Sometimes there is a small anal sinus. At the base of the shell, the distal end of the short, oblique, widely-open siphonal canal meets the palatal lip at a slightly-acute or right angle when viewed end-on 13 Tritia reticulata & 14 Tritia reticulata . The short, straight, white, exposed columella forms one side of the canal. The siphonal canal of earlier stages remains as a deep groove on the columella concealed within the spire for its entire length to the apex of the shell 15 Tritia reticulata . The substantial, opaque, glossy white, semicircular parietal lip spreads extensively over the body whorl 12 Tritia reticulata . It may be stained by epizooic growth 16 Tritia reticulata . The porcelaneous (when live or recently dead) white aperture interior rapidly thickens away from the thin palatal rim and, on adults, forms a rib with 6 to 10 unevenly sized palatal teeth. The one or two near the middle are often the most prominent 12 Tritia reticulata . Frequently there are up to six less conspicuous columellar/parietal teeth 17 Tritia reticulata . The external ground colour of the shell is brownish when the periostracum is intact, but opaque, matt whitish/buff when the periostracum is eroded 07 Tritia reticulata . Exposed shell may be stained by epizoic growth 16 Tritia reticulata . Spire whorls may have a brown or purple-brown subsutural line, and the body whorl may have a similar line on its periphery and another above the spiral gutter18 Tritia reticulata . The small, oval, translucent, horn-coloured operculum19 Tritia reticulata has a distal (outer) surface of conchiolinwith concentric growth rings round a peripheral nucleus. The proximal (inner) surface has a matt surface where the opercular disc attaches to it, with concentric lines running transversely to exterior growth lines. The unattached part of the proximal surface is of varnish without growth lines.
Body description
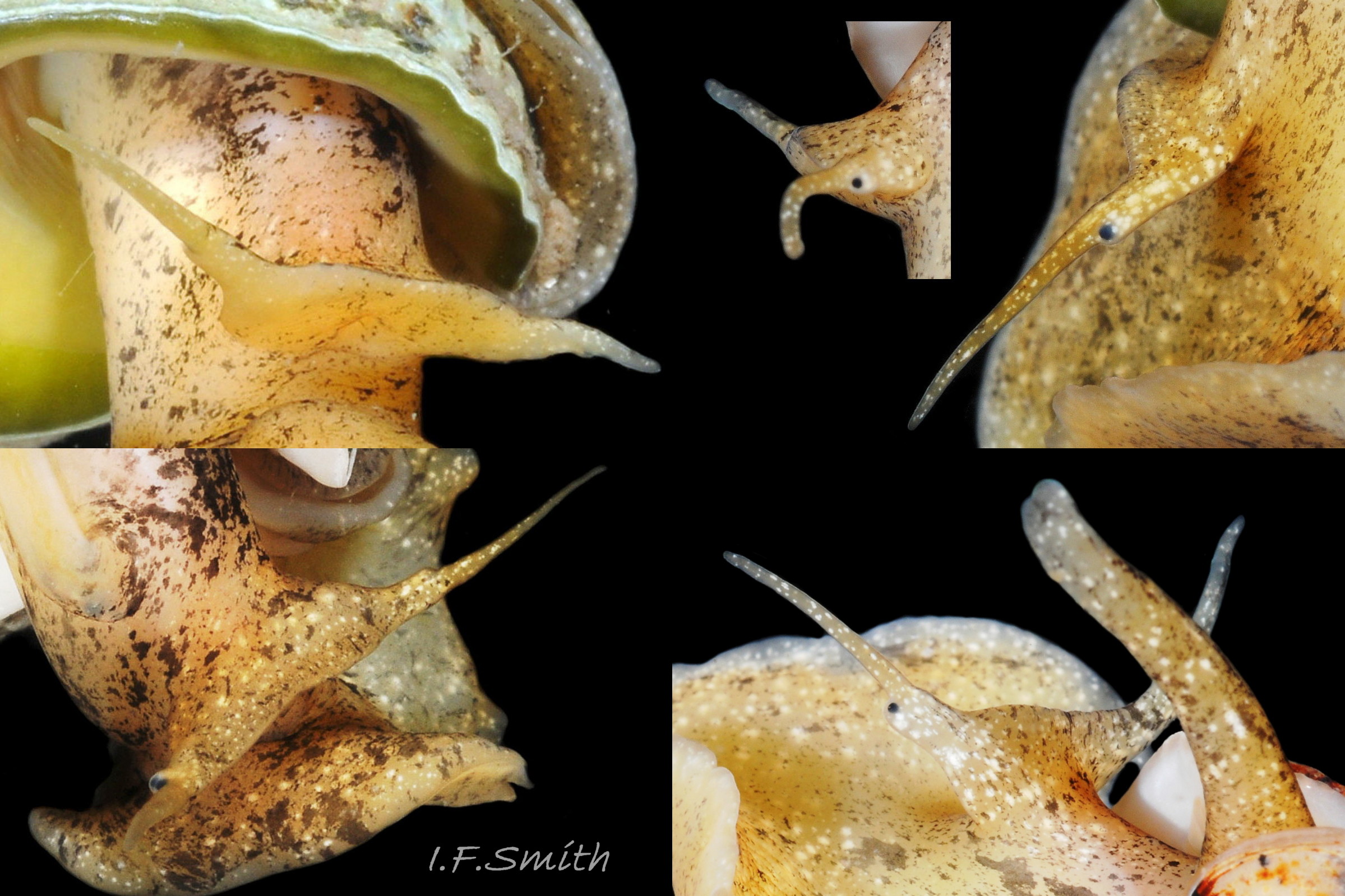
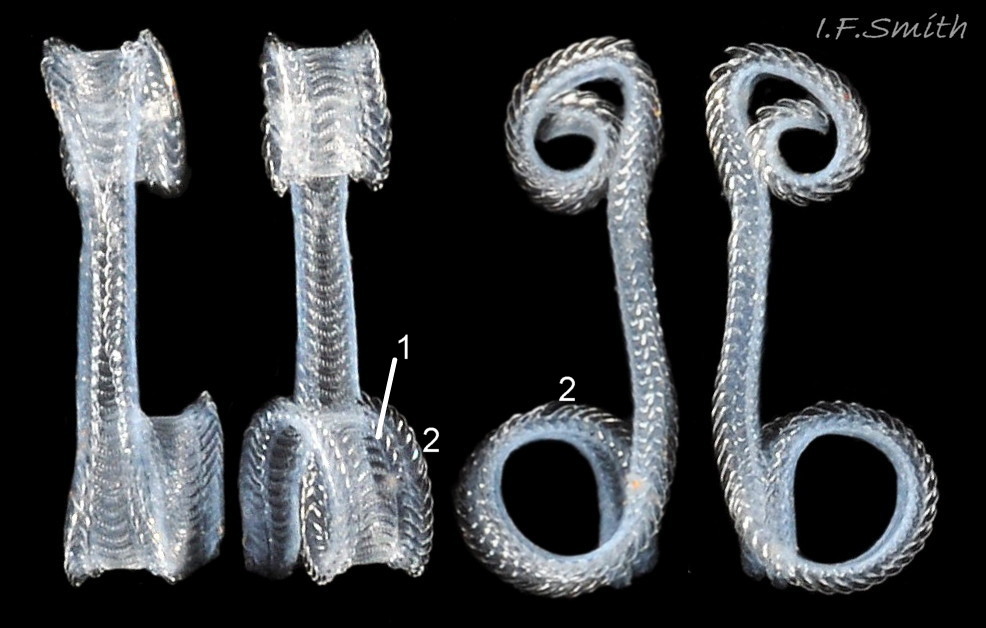
The ground colour of most parts of the body is usually light sienna or yellowish 11 Tritia reticulata or, on juveniles, whitish 20 Tritia reticulata, heavily marked with small white and brown/black marks. It is uncertain whether grey bodies are confined to T. nitida, as stated by Jeffreys (1867); they might occur on both spp. The rectangular head is dorsoventrally flattened with a long, slender, tapering cephalic tentacle on each anterior corner 21 Tritia reticulata. There is a small, black eye on the swollen base of each tentacle. There is no external snout between the tentacles, but the ventral surface of the head has a slit opening (pseudo-mouth) 22 Tritia reticulata of a sac containing the retracted, long, unmottled, pleurembolic 23 Tritia reticulata proboscis 24 Tritia reticulata & 25 Tritia reticulata . The radula, buccal mass and the true mouth are in the distal end of the proboscis. The rachiglossan radula has in each row of colourless, transparent teeth a wide, central, rachidian tooth with many small cusps (points) flanked on either side by a longer marginal tooth 26 Tritia reticulata . The yellow mantle lacks mottling at the anterior of the mantle cavity 16 Tritia reticulata , but it has much dark pigment further back 27 Tritia reticulata. The mantle on the left extends as contractile, densely mottled, respiratory siphon through, and often far beyond, the siphonal canal of the shell 28 Tritia reticulata . The mantle can extend widely onto the body whorl exterior to deposit the wide, glossy white parietal lip 16 Tritia reticulata . Within the mantle cavity, behind the right tentacle, males have a large, curved, buff-white penis with only a few dark marks basally 29 Tritia reticulata & 30 Tritia reticulata . Also in the mantle cavity (dissection required) are a large, yellow-brown, pinnate ctenidium and a large, pinnate, red-brown osphradium resembling a ctenidium in structure 31 Tritia reticulata .
The large, broad, ovoid foot 32 Tritia reticulata is truncate at the anterior with angulated corners (propodial tentacles) that can be varied from bluntly rounded 33 Tritia reticulata to recurved and sharply pointed 34 Tritia reticulata . The anterior and corners of the foot are deeply bilaminate 35 Tritia reticulata with the anterior pedal mucus gland within. In life, the bilaminate groove is often concealed by the upper lamina overhanging it. The posterior of the foot narrows to a blunt point with a pair of small metapodial tentacles dorsally 05 Tritia reticulata . The foot can transform to a variety of shapes, and twist abruptly to different positions 36 Tritia reticulata . Dorsally the foot is coloured as most of the body 32 Tritia reticulata but on the sole white marks are usually limited to near the edge 06 Tritia reticulata .
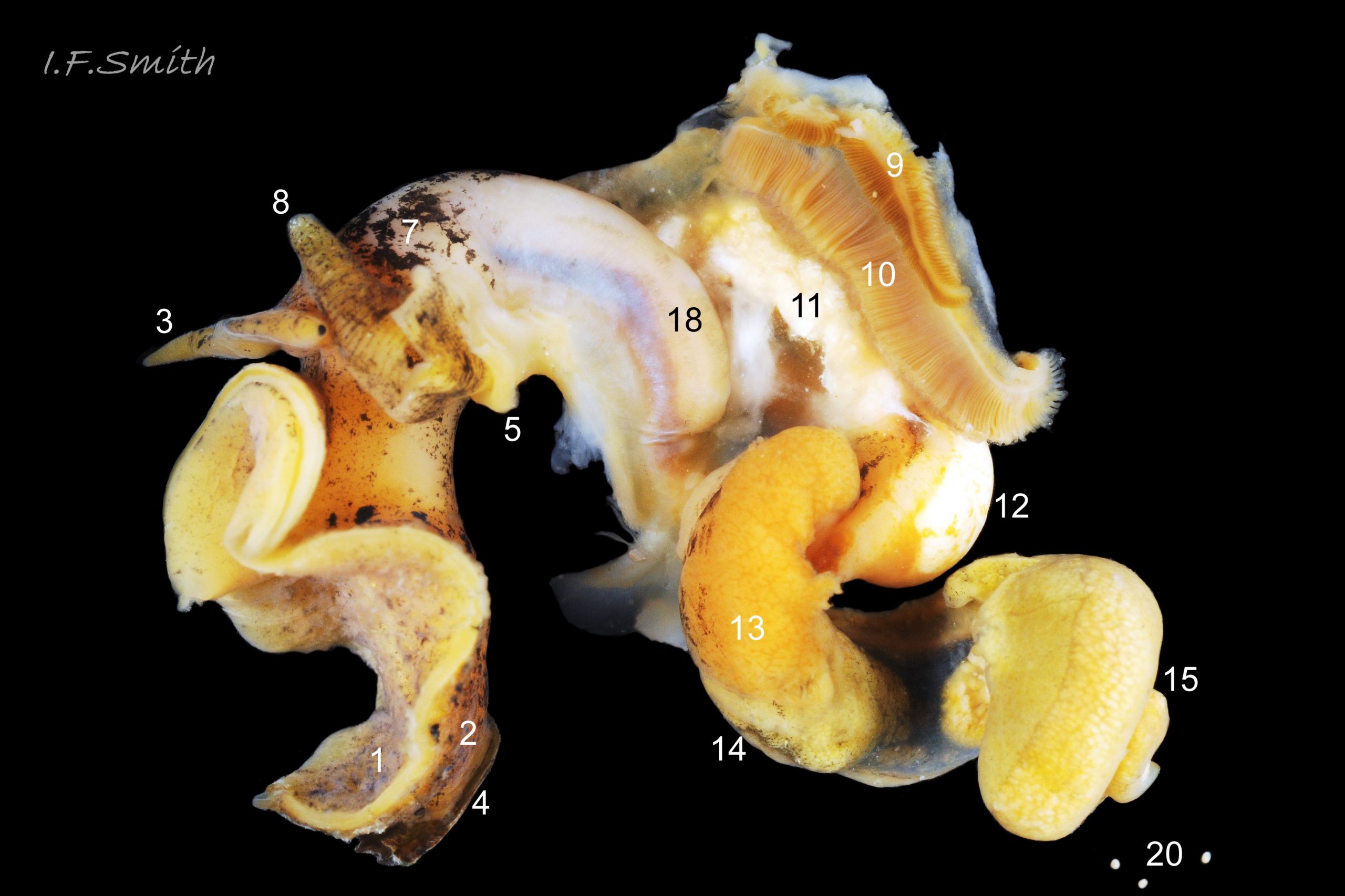
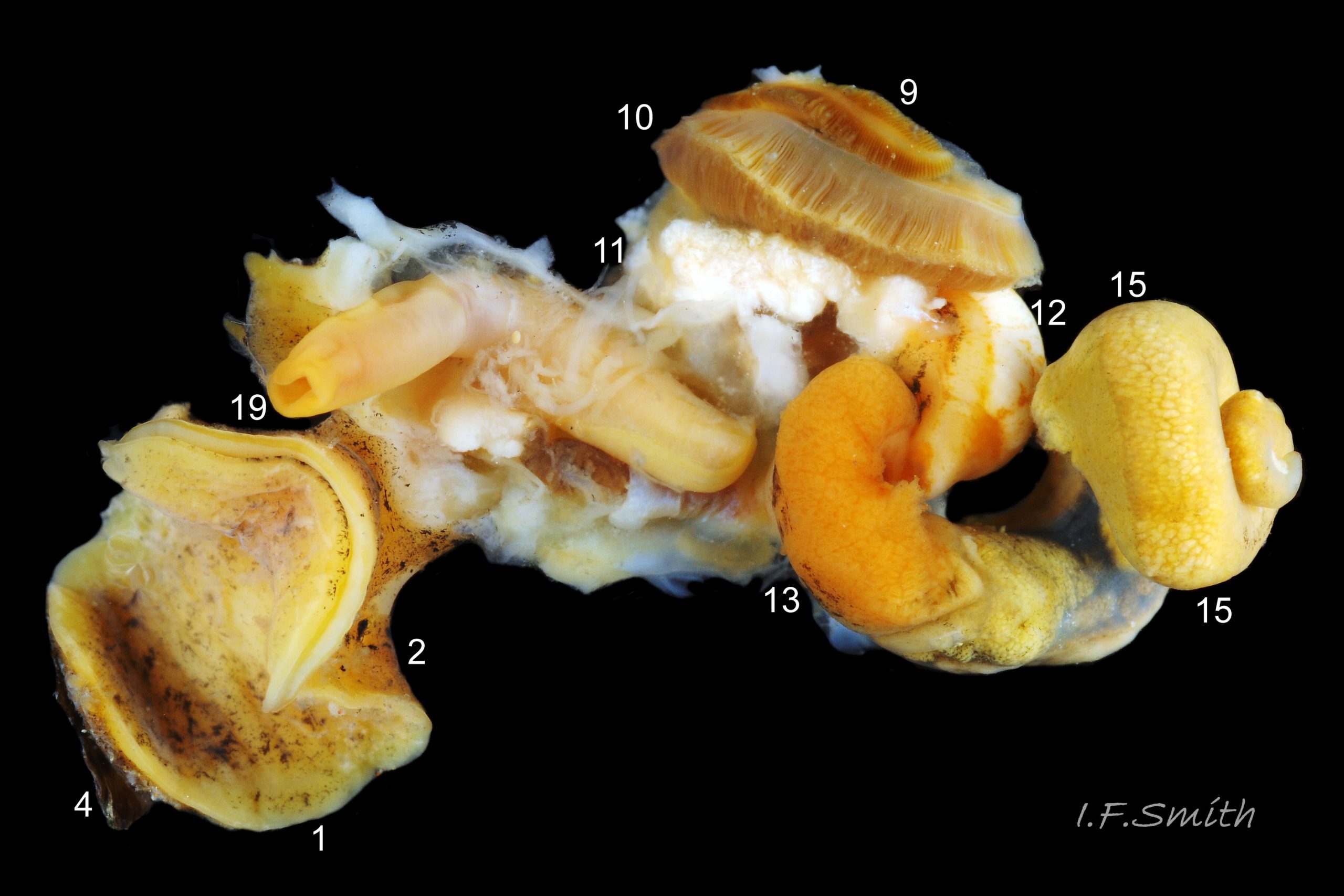
The dissected female in images 27 Tritia reticulata (mantle intact) , 24 Tritia reticulata (mantle cut and opened to right, but remaining on part of the mantle cavity) and 25 Tritia reticulata (body wall opened) , shows the following features:
1: sole of foot. 2: dorsum of foot. 3: cephalic tentacle.
4: operculum. 5: mantle edge. 6: unblotched mantle.
7: darkly blotched mantle. 8: respiratory inhalent siphon.
9: osphradium for testing inhalent water. 10: ctenidium (gill).
11: hypobranchial gland, secretes mucus to trap particles from inhalent water.
12: kidney. 13: digestive gland. 14: stomach. 15: ovary.
16: auricle of heart. 17: ventricle of heart.
18: proboscis sheath seen through translucent body wall.
19: proboscis with sheath removed. 20: ova.
Key identification features
Identification should be made using a combination of features as some elements may vary.
Tritia reticulata
1) Maximum height 30 mm.
2) Spire variably slender, flat-sided, sharply pointed cone with almost flat whorls on spire and shallow suture 01 Tritia reticulata (Body whorl convex). Juveniles are squatter, taper more sharply and may have a slightly concave-sided spire 02 Tritia reticulata.
3) Smooth protoconch, c.1 mm diameter, usually persists to form a fine, sharp apex 08 Tritia reticulata , even when worn 10 Tritia reticulata .
4) Costae (axial ribs) intersect with spiral ridges to create reticulate arrangement of squarish bosses 07 Tritia reticulata . In Britain, number of costae on body whorl is 15 to 23 (pers. obs. IFS), (15 to 20 in Jeffreys, 1867) and, in Ria de Vigo, Spain, 16 to 23 (Rolan and Luque, 1994).
5) Only very rarely, is the final costa sufficiently thickened to form a slight labial varix 04 Tritia reticulata .
6) Teeth within the outer (palatal) lip are unevenly sized, one or two near middle often most prominent 12 Tritia reticulata . The inner (columellar and parietal) lip is “more or less tuberculated” (Jeffreys, 1867), but teeth/small ribs may be absent, especially on young ones.
7) When live and unworn, a brown periostracum covers the shell except the protoconch and white rim of outer (palatal) lip 05 Tritia reticulata . When the periostracum is removed, the shell exterior is matt whitish/buff 07 Tritia reticulata , unless stained by epizooic growth or other matter 16 Tritia reticulata . Interior is glossy white 03 Tritia reticulata .
8) Semicircular, glossy, opaque, white parietal lip covers large part of body whorl 12 Tritia reticulata ; may be stained by epizooic growth or other matter 16 Tritia reticulata .
9) White columella and siphonal canal. No dark spot within canal 03 Tritia reticulata .
10) Siphonal canal meets palatal lip at slightly-acute or right angle when viewed end-on 13 Tritia reticulata . Beachworn specimens may have angle worn to obtuse and resemble T. nitida 37 Tritia reticulata .
11) 4 to 5 grooves on columellar neck of siphonal canal 07 Tritia reticulata [Often worn away, so use 4 or 5 to confirm T. reticulata; but those with 3, fewer or none may be either T. nitida or worn T. reticulata 11 Tritia reticulata & 38 Tritia reticulata .]
12) Body flesh light sienna/yellowish 11 Tritia reticulata (juveniles, whitish 20 Tritia reticulata), heavily marked with small blotches of white and brown/black. Uncertain whether grey body only on T. nitida.
13) Egg capsules like a flat, circular brandy bottle that broadens with age 39 Tritia reticulata . Mean height 4.07 mm, width 3.25 mm, W/H 80% including neck (Rolan & Luque, 1994).
14) Sandy sediment sublittorally, and LWS on rocky shores with areas of sand, all round Britain and Ireland.
Similar species
Tritia nitida (Jeffreys, 1867)
WoRMS accepts T. nitida as a valid species; supported by chromotology (Collyer, 1961), comparative morphology (Rolan & Luque, 1994), allozymes (Sanjuan et al., 1997) and DNA (Couceiro et al., 2012); further detail in Smith (2019). Populations vary geographically, so some features may not match those below. They are marked (J) for Thames, Orwell and Roach estuaries in S.E. England in Jeffreys (1867); (B) for a sample of 220 in 2019 from Blackwater estuary in S.E. England; (S) for Ria de Vigo, N.W. Spain in Sanjuan et al. (1997); (R) for Ria de Vigo in Rolan and Luque (1994) and (P) for northern Adriatic Italy and Croatia (J. Prkić, 2019, pers. comm. September 2019). So use a combination of features when making an identification.
1) Maximum height 25 mm (J), exceptionally (2% of strandline sample) 26 to 30 mm (B) 46 Tritia reticulata , 30 mm in Galicia, Spain (Trigo et al., 2018) and 40 mm in the Italian Adriatic (P) 47 Tritia reticulata .
2) AdultT. nitida spire-whorls convex in Essex46 Tritia reticulata & 48 Tritia reticulata . Jeffreys (1867), stated “whorls flattened; – – suture deeper [than on T. reticulata]”. Compared to gastropods generally, the gentle convexity of the whorls on Essex T. nitidacould be said to be flattish, but they are more convex than on T. reticulata, resulting in the deeper suture Jeffreys mentions. In the Adriatic, T. nitida whorls vary from convex to nearly flat 50 Tritia reticulata .
3) Smooth protoconch is frequently worn/broken off to give a blunt apex 48 Tritia reticulata but it appears to persist more on Mediterranean T. nitida 50 Tritia reticulata .
4) In Britain, number of costae on body whorl is 10 to 14 B 56 Tritia reticulata , [10 to 12, J]
In southern Europe T. nitida varies more widely, 11 to 19 (R), 9 to 18 (P), and overlaps with T. reticulata when 15 or more costae.
5) Labial varix on almost all shells over 15 mm high (J, B, P) and additional varix on body whorl occasionally in Britain (J) 51 Tritia reticulata , and frequently in Adriatic (P) 50 Tritia reticulata . In Britain the varix is usually less pronounced than on T. incrassata; sometimes little more than slightly thickened costae, and lost or inconspicuous on some beachworn strandline shells 52 Tritia reticulata . Adriatic specimens can have very prominent varices 53 Tritia reticulata & 54 Tritia reticulata .
6) Teeth within the outer (palatal) lip quite evenly sized 55 Tritia reticulata , sometimes one a bit higher. “inner [columellar/parietal] lip – – – never tuberculated” (J), but some Adriatic T. nitida have tubercles/teeth on the inner lip 55 Tritia reticulata .
7) “epidermis [periostracum] inconspicuous 57 Tritia reticulata , or obscured by an earthy incrustation 58 Tritia reticulata ” (Jeffreys, 1867). Shell exterior and interior 59 Tritia reticulata violaceous (R&S), purplish on yellowish white ground, raised sculpture yellowish white, and coloured lines 57 Tritia reticulata brighter purple than on T. reticulata (J) but presence and intensity of purple varies greatly, often absent or, on beachworn shells, faded (B) 60 Tritia reticulata .
8) In Britain, parietal lip translucent showing shell colour beneath (S & R) 48 Tritia reticulata ; not as extensive, white or distinct as on T. reticulata, but strong and opaque on some T. nitida in Adriatic 53 Tritia reticulata & 47 Tritia reticulata . Sometimes eroded or stained on beachworn shells 48 Tritia reticulata .
10) Siphonal canal meets palatal lip at obtuse angle when viewed end-on (R) 61 Tritia reticulata . [Beachworn T. reticulata may have angle worn to obtuse and resemble T. nitida 52 Tritia reticulata .]
11) Three grooves on columellar neck of siphonal canal 62 Tritia reticulata but eroded from most strandline shells (75% B) and some live shells.
12)Jeffreys (1867) described the body of T. nitida as “greyish, with a slight tinge of purple, and closely speckled with flake-white”. This fits the Bretagne specimen at 63 Tritia reticulata , presumably those from Essex where Jeffreys took his specimens, and some from the Adriatic 58 Tritia reticulata , but some other Adriatic T. nitida have yellowish bodies 64 Tritia reticulata .The respiratory siphon is “almost always blackish” in Ria de Vigo (R) but both blackish and paler siphons occur on Adriatic T. nitida 58 Tritia reticulata & 64 Tritia reticulata . The value of flesh and siphon colours for distinguishing T. nitida from T. reticulata elsewhere is uncertain.
13) T. nitida egg capsules are slenderly ovoid (R&S) with W/H 65% including neck 39 Tritia reticulata & 65 Tritia reticulata . Mean height 2.65 mm, width 1.72mm (R) W/H 65%.
14) Sublittoral on muddy substrate in sheltered, variable salinity on Atlantic coasts (S & J). Scattered sites Spain to S. Sweden; more continuous distribution in Mediterranean beyond Barcelona and in Black Sea at variable, often high, salinity (S & R).
Tritia incrassata (Strøm, 1768)
1) Maximum height 12 mm.
2) Squatter shell than T. reticulata, adult whorls convex 66 Tritia reticulata.
3) Strong, smooth protoconch has c.3 whorls, diameter 0.5 to 0.6 mm (Fretter & Graham, 1985), distinctly demarcated from teleoconch. Usually intact, or with only minor damage, when live.
4) Shell sculpture of transverse, oblong, rather than squarish, bosses 66 Tritia reticulata.
5) Pronounced, broad, labial varix; light brown when covered by periostracum 66 Tritia reticulata; white, often with transverse red-brown bands when periostracum worn away 67 Tritia reticulata.
7) Periostracum only retained in grooves of sculpture on live specimens 66 Tritia reticulata.
8) Semicircular, glossy, opaque, white parietal lip covers part of body whorl, less extensive than on T. reticulata 66 Tritia reticulata.
9) White columella and siphonal canal 66 Tritia reticulata, with distinct brown or black spot inside canal 67 Tritia reticulata (often hidden in photographs by shadow, the siphon or the tilt of the pose).
13) Egg capsules small, 1.5 mm to 2 mm high, similar form to that of T. reticulata, but with longer neck, narrow stalk-base and laid in irregular clumps 68 Tritia reticulata (image in Lebour, 1931, http://plymsea.ac.uk/id/eprint/697/ ).
14) Rocky shores all round Britain. Lower shore and sublittoral.
Tritia varicosa (W. Turton, 1825)
1) Maximum height 14 mm.
2) Squatter shell than T. reticulata, adult whorls convex 69 Tritia reticulata .
3) Smooth protoconch has 2.25 whorls , diameter 750 to 950µ (Fretter & Graham, 1985), nearly as large as on the much larger shell of T. reticulata 70 Tritia reticulata .
4) Widely-spaced, narrow costae intersect with narrowly-spaced, narrow spiral ridges to form small, rounded bosses 69 Tritia reticulata . All sculpture is well-raisedon unweathered shells.
5) Pronounced labial varix on adult shells, and most have one or more additional varices on body-whorl or spire whorls. Varices usually white with transverse brown bands 69 Tritia reticulata .
7) Shell glossy with no obvious periostracum when live.
9) Columellar lip orange-brown. Often orange-brown at entrance of siphonal canal, but no black or blackish brown further within canal 70 Tritia reticulata.
10) Viewed end on, rounded siphonal canal meets palatal lip at an acute angle 70 Tritia reticulata .
14) South and west coast of Britain to Shetland. Not Irish Sea. Sublittoral, 1 m to 200 m, sandy substrate; found on bait in crab pots.
Habits and ecology
T. reticulata lives in full marine salinity at LWS on shores with mixed hard and sandy substrate at spots where organic remains accumulate, and on sandy substrate to about 35 metres sublittorally. Large congregations may occur on carrion 40 Tritia reticulata . Reports of it in brackish water >16‰ on muddy substrate may be of unrecognized T. nitida, which also lives at high salinity in the Mediterranean. At low tide on shores, pools and runnels are favoured or shelter is taken under rocks or below wet sediment with the respiratory siphon protruding. The siphon tip may have photoreceptors as it withdraws when shadow falls on it.
Sublittorally, it remains hidden in the surface layer of sediment with its inhalent siphon tip protruding for much of the time, emerging promptly when carrion or faeces are sensed by its large osphradium and gliding rapidly to it on its broad foot lubricated with mucus from the anterior pedal gland. Many assemble like vultures 49 Tritia reticulata and push their proboscises, 150% length of shell at full extension, deep into carrion while holding their long siphons away from it to inhale uncontaminated water 40 Tritia reticulata . The sucker-like tip enables the proboscis to retain feeding position while the radula rasps 25 Tritia reticulata . The faeces are oval pellets.
The strong shells of adults resist crab attack, but many beached, thin, juvenile shells have broken lips which may indicate successful crab predation 41 Tritia reticulata . Predatory Euspira spp. may bore the shells of juvenile T. reticulata near the apex where thinnest, but below the persistent protoconch 42 Tritia reticulata &15 Tritia reticulata . When starfish attack, T. reticulata reacts immediately to first touch by vigorously twisting and turning its body and shell in different directions to confuse the attacker 36 Tritia reticulata .
It breeds from March to August at Plymouth, and from April to August further east in the English Channel (Fretter & Graham, 1985). The female inspects potential ovipositing sites of rock, shell, Zostera or alga; with the tip of her siphon, and may clean the surface with its radula before laying capsules containing 50 to 350 orange-pink 43 Tritia reticulata or white 44 Tritia reticulata ova in each. The capsule passes along a temporary groove in the foot to the female ventral pedal gland 36 Tritia reticulata which grips it and forms it into a flattened, brandy-bottle shape with a circular chamber, flat on one face and slightly convex on the other. The small base is secured in position on a large disc of cement. The mean capsule size, including apical neck, is 4.07 mm high and 3.25 mm width W/H 80% (Rolan & Luque, 1994). The capsule and fluid around the ova are transparent colourless. Initially, ova fill less than half of the capsule but by the end of one to two months’ development the capsule has broadened c.20% and ova have grown to fill it 39 Tritia reticulata . Capsules are often laid in rows like books on a shelf all leaning at c.50° 43 Tritia reticulata . Hatched, free-swimming veligers escape through the now unplugged apical neck of the capsule. Veligers have a transparent, smooth shell of 1.5 whorls, c. 300µlong. The bilobed velum has a marginal brown line, and the sole is red-brown. After two to three 3 months in the plankton (mainly June to September, with a few January to March, at Plymouth), it metamorphoses to a three whorled, c.750µ, shell with siphonal canal (Fretter & Graham, 1985; may include data of T. nitida).
Distribution and status
T. reticulata sensu stricto lives along Atlantic coasts from Norway to Morocco and into the Mediterranean only as far east as Oran, Algeria and 23km north-east of Barcelona, being replaced further east and into the Black Sea by T. nitida form mamillata (Rolan and Luque, 1994). Images of T. reticulata sensu stricto on iNaturalist at https://www.inaturalist.org/observations/168276172 show that it occurs further east to near Marseille, France. The Norwegian marine gastropod list by Høisæter (2009) omits T. reticulata sensu stricto, but images from Norway by E. Svensen show that it is present 45 Tritia reticulata & 40 Tritia reticulata . The GBIF map https://www.gbif.org/species/5727892 for T. reticulata should be regarded as showing distribution of an aggregate with T. nitida. T. reticulata</i > is common all round Britain and Ireland, except scarce or absent from Fife to Caithness, and there are few records from Suffolk, Lincolnshire and Cumbria. Those in sheltered muddy estuaries in Essex need careful examination as T. nitida occurs commonly there, and possibly in other sheltered waters. U.K. interactive map NBN (aggregated with T. nitida) https://records.nbnatlas.org/occurrences/search?q=lsid:NHMSYS0021185422#tab_mapView
Acknowledgements
I am indebted to Philippe Boissel, Carlos Fernández-Cid, Joe FitzGibbon, Aleksander Golemaj, Alen Petani, Bas van der Sanden, Erling Svensen, Mark Thomas, Pero Ugarković, Stefan Verheyen and Neil Ward for use of their informative images, to Miquel Pontes for providing vital literature, and to Julia Nunn, Jakov Prkić and Vollrath Wiese for information and advice. Special thanks are due to my co-author, Simon Taylor, for undertaking shore visits and supplying specimens for this account, and to Jakov Prkić for taking many images specifically for this account.
Links and references
Chaster, G.W., Knight, G.A.F., Melvill, J.C. and Hoyle, W.E. 1901. List of British marine mollusca and brachiopoda. Manchester, Conchological Society of Great Britain and Ireland. https://archive.org/details/listofbritishmol00chas/page/n21/mode/2up
Collins, K.S., Edie, S.M., Gao, T., Bieler, R. and Jablonski, D. 2019. Spatial filters of function and phylogeny determine morphological disparity with latitude. PLoSONE 14 (8): e0221490. https://doi.org/10.1371/journal.pone.0221490
Collyer, D.M. 1961. Differences revealed by paper partition chromatography between the gastropod Nassarius reticulatus (L.) and specimens believed to be N. nitida (Jeffreys). J. Mar. Biol. Ass. 41(3): 683 to 693.
Couceiro, L., López, L., Sotka, E.E., Ruiz, J.M. & Barreiro, R. 2012. Molecular data delineate cryptic Nassarius species and characterize spatial genetic structure of N. nitidus. J. Mar. Biol. Ass. 92(5): 1175 to 1182. https://www.researchgate.net/publication/230846249_Molecular_data_delineate_cryptic_Nassarius_species_and_characterize_spatial_genetic_structure_of_N_nitidus
Diz, P., Jorissen F. J., Reichart, G. J., Poulain, C., Dehairs, F., Leorri E. and Paulet, Y.-M.
Interpretation of benthic foraminiferal stable isotopes in subtidal estuarine environments. Biogeosciences, 6, 2549–2560, 2009 www.biogeosciences.net/6/2549/2009/
Forbes, E. & Hanley S. 1849-53. A history of the British mollusca and their shells. vol. 3 (1853), London, van Voorst. (As Nassa reticulata) https://archive.org/stream/historyofbritish03forbe#page/388/mode/2up/search/reticulatus
Fretter, V. and Graham, A. 1985. The prosobranch molluscs of Britain and Denmark. Part 8 – Neogastropoda. Suppl. 15, J. Moll. Stud.
Graham, A. 1988. Molluscs: prosobranch and pyramidellid gastropods. Synopses of the British Fauna (New Series) no.2 (Second edition). Leiden, E.J.Brill/Dr. W. Backhuys. 662 pages.
Hayward, P.J. & Ryland, J.S. (eds.) 1995. Handbook of the marine fauna of North-West Europe. Oxford University Press.
Høisæter, T. (2009). Distribution of marine, benthic, shell bearing gastropods along the Norwegian coast. Fauna Norvegica, 28: 5-106. https://doi.org/10.5324/fn.v28i0.563
Jeffreys, J.G. 1862-69. British conchology. vol. 4 (1867). London, van Voorst. (As Nassa reticulata) p346 in pdf https://archive.org/stream/britishconcholog04jeffr#page/346/mode/2up .
Original description of T. nitida (as Nassa nitida) on p349 at https://archive.org/stream/britishconcholog04jeffr#page/348/mode/2up
Lebour, M. V. 1931. The larval stages of Nassarius reticulatus and Nassarius incrassatus. J. Mar. Biol. Ass., 17 (3). 797-818. http://plymsea.ac.uk/id/eprint/697/
Linnaeus, C. (1758). Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata [10th revised edition], vol. 1: 824 pp. Laurentius Salvius: Holmiae. https://www.biodiversitylibrary.org/page/726886#page/762/mode/1up entry 411 p. 740
Marshall, J. T. 1895. Alterations in ‘British Conchology’. J. Conch. 8: 24 to 41.
https://www.biodiversitylibrary.org/item/99811#page/60/mode/1up [p38 as Nassa nitida]
McKay, D.W. & Smith, S.M. 1979. Marine Mollusca of East Scotland. Edinburgh, Royal Scottish Museum.
McClelland, H. 1926. General index of all families, genera, species and varieties described and noted in the Journal of Conchology vols. I – XVI, 1874 to 1922. Proceedings of the Malacological Society of London vols. I – XV, 1893 to 1923. The Conchologist vols I – II, 1891 to 1893, continued as the Journal of Malacology vols. III – XII, 1894 to 1905. Birmingham, Birbeck and sons.
McMillan, N.F. 1968. British shells. London, F.Warn.
Rolan, E. and Luque, A.A. 1994. Nassarius reticulatus (Linnaeus, 1758) and Nassarius nitidus (Jeffreys, 1867) (Gastropoda, Nassariidae), dos especies válidas de los mares de
Europa. Iberus, 12 (2): 59-76. https://www.biodiversitylibrary.org/part/98444
Sanjuan, A., Pérez-Losada, M. & Rolan, E. 1997. Allozyme evidence for cryptic speciation in sympatric populations of Nassarius spp. (Mollusca: Gastropoda). J. Mar. Biol. Ass. 77(3): 773 to 784.
Smith, I.F. 2019. Differentiation of Tritia reticulata (Jeffreys, 1867) from Tritia reticulata (Linnaeus, 1758). Mollusc World Issue 51. Expanded pdf version at https://www.researchgate.net/publication/336441072_Recognition_of_Tritia_nitida_in_Britain_and_Ireland
Trigo, J.E.; Diaz Agras, G.J.; Garcia Alvarez, O.L.; Guerra, A.; Moreira, J.; Pérez, J.; Rolán, E.; Troncoso, J.S,; Urgorri, V.. 2018. Guia de los Moluscos Marinos de Galicia. Servicio de Publicacións da Universidade de Vigo.
Turton, W. 1825. Description of some new British shells. Zoological Journal, 2: 361-367. [Fist description of Tritia varicosa as Tritonia varicosa]. https://www.biodiversitylibrary.org/page/2255625#page/431/mode/1up
Yonge, C.M. and Thompson, T.E. 1976. Living marine molluscs. Collins, London. [Fig. 52G shows Danish egg capsules of T. nitida labelled as Nassarius reticulatus.]
Glossary
acrembolic = (of proboscis) introversible/eversible like finger of glove.
adapical = towards the apex of the shell.
anal sinus = notch in adapical angle of aperture; route of rectum/anus to exterior of shell.
aperture = mouth of gastropod shell; outlet for head and foot.
bilaminate = (adj.) formed of two layers.
cephalic = (adj.) of or on the head.
cilia = (pl.) vibrating linear extensions of membrane used in locomotion..
ciliated = (adj.) coated with cilia.
coll. = (or “in coll.”, abbreviation of “in collectionem”) in the collection of (named person or institution; c.f. leg.).
columella = axis of gastropod shell spiral, exposed on final whorl by aperture.
columellar = (adj.) of or near central axis of spiral gastropod.
columellar lip = lower (abapical) part of inner lip of aperture.
costa = (pl. costae) axial rib crossing shell whorl at about right-angles to any spiral striae.
costal = (adj.) of, or arranged like, costae.
ctenidium = comb-like molluscan gill; usually an axis with filaments either side.
cusp = raised point or prominence on crown of a tooth.
distal = away from centre of body or from point of attachment.
dorsoventrally flattened = as if pressed flat from above.
ELWS = extreme low water spring tide (usually near March and September equinoxes).
height = (of gastropod shells) distance from apex of spire to base of aperture.
hypobranchial gland = thickened, sometimes puckered, tissue on roof of mantle cavity of many gastropods. Secretes mucus to trap and consolidate particles from inhalent water. Often other biologically active compounds produced.
labial varix = especially strong or broad costa (rib) along edge of outer lip of aperture.
leg. = (abbreviation of legit) collected/ found by (c.f. coll.)
LWS = low water spring tide, two periods of a few days each month when tide falls lowest.
mantle = sheet of tissue that secretes shell and forms a cavity for the gill in most marine molluscs.
metapodial = (adj.) of the hind part of the foot.
odontophore = tongue-like structure of cartilage supporting radula.
opercular = (adj.) of the operculum.
operculum = plate of horny conchiolin, rarely calcareous, used to close shell aperture.
osphradium = organ for testing water for pollutant, prey or predator chemicals and/or for particles.
periphery = ‘equator’ of gastropod body whorl, sometimes with a keel or coloured band.
periostracum = thin horny layer of chitinous material often coating shells.
pers. comm. = personal communication by face-to-face conversation, telephone, letter or email.
plankton = animals and plants that drift in pelagic zone (main body of water).
pleurembolic proboscis = basal part (only) of proboscis inverts to form a sac for rest of proboscis to be retracted into without inversion. (c.f. acrembolic).
porcelaneous = resembling vitreous glazed ceramic material.
protoconch = apical whorls produced during embryonic and larval stages; different in form from other whorls forming teleoconch.
proximal = towards the centre of the body or point of attachment.
rachidian = (adj.) median/middle tooth in each row of teeth on radula.
rachiglossan = (adj.) of radula with a many-cusped, rachidian tooth, and single marginal tooth at each side.
radula = chitinous ribbon of teeth; extended on odontophore to rasp food.
sigmoid = curved in two directions like letter ‘S’ but often with shallower curvature.
subsutural = close below the suture when shell positioned with apex uppermost.
sutural = (adj.) of or close to a suture.
suture = groove or line where whorls of gastropod shell adjoin.
teleoconch = entire gastropod shell, apart from apical protoconch.
varix = (see labial)
veliger = shelled larva of marine mollusc; swims by waving cilia on velum (bilobed flap).