Click image to enlarge with full caption. Main text below slider.
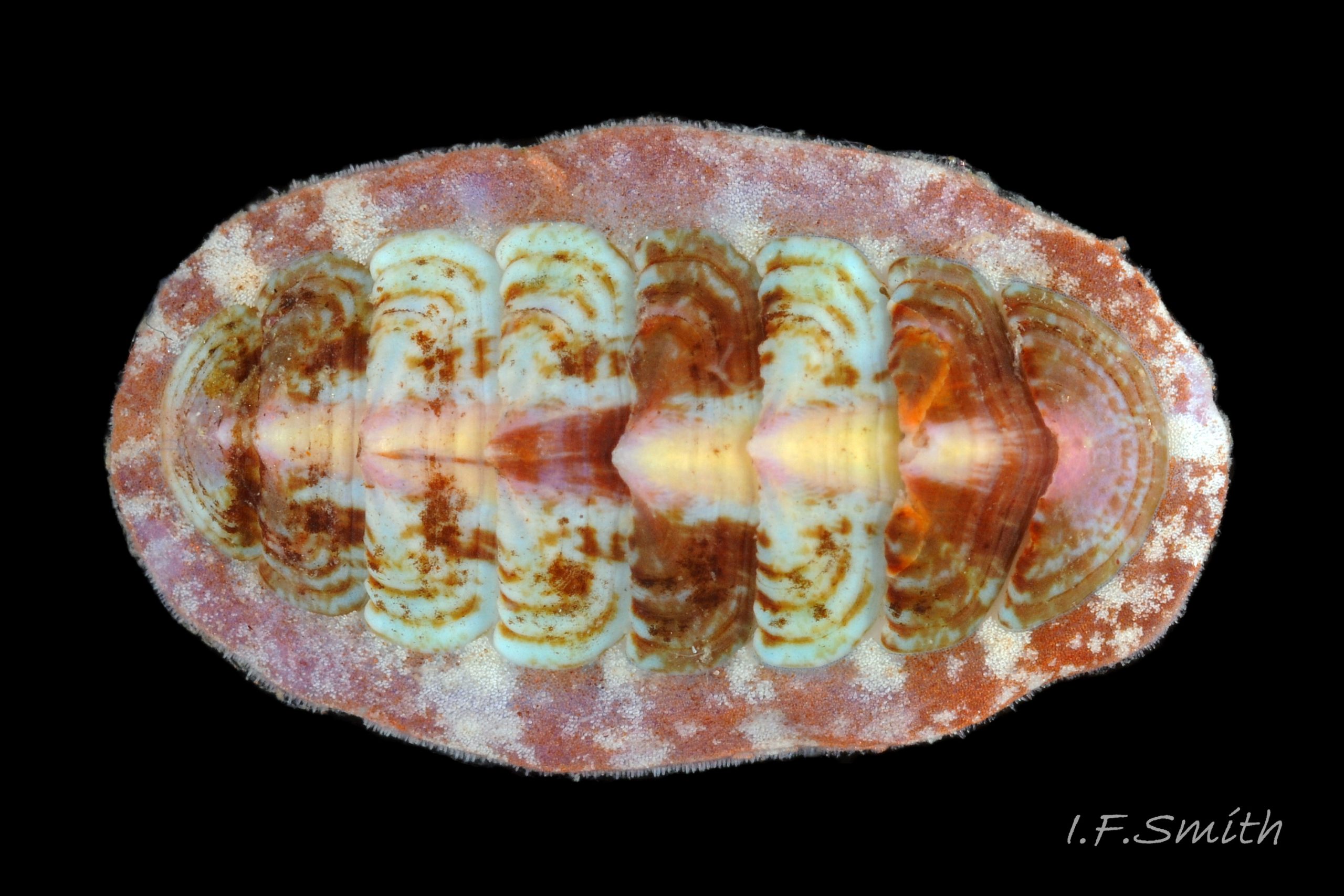
01 Boreochiton ruber. Length 12mm. North Yorkshire. September 2014. (leg. Paula Lightfoot).
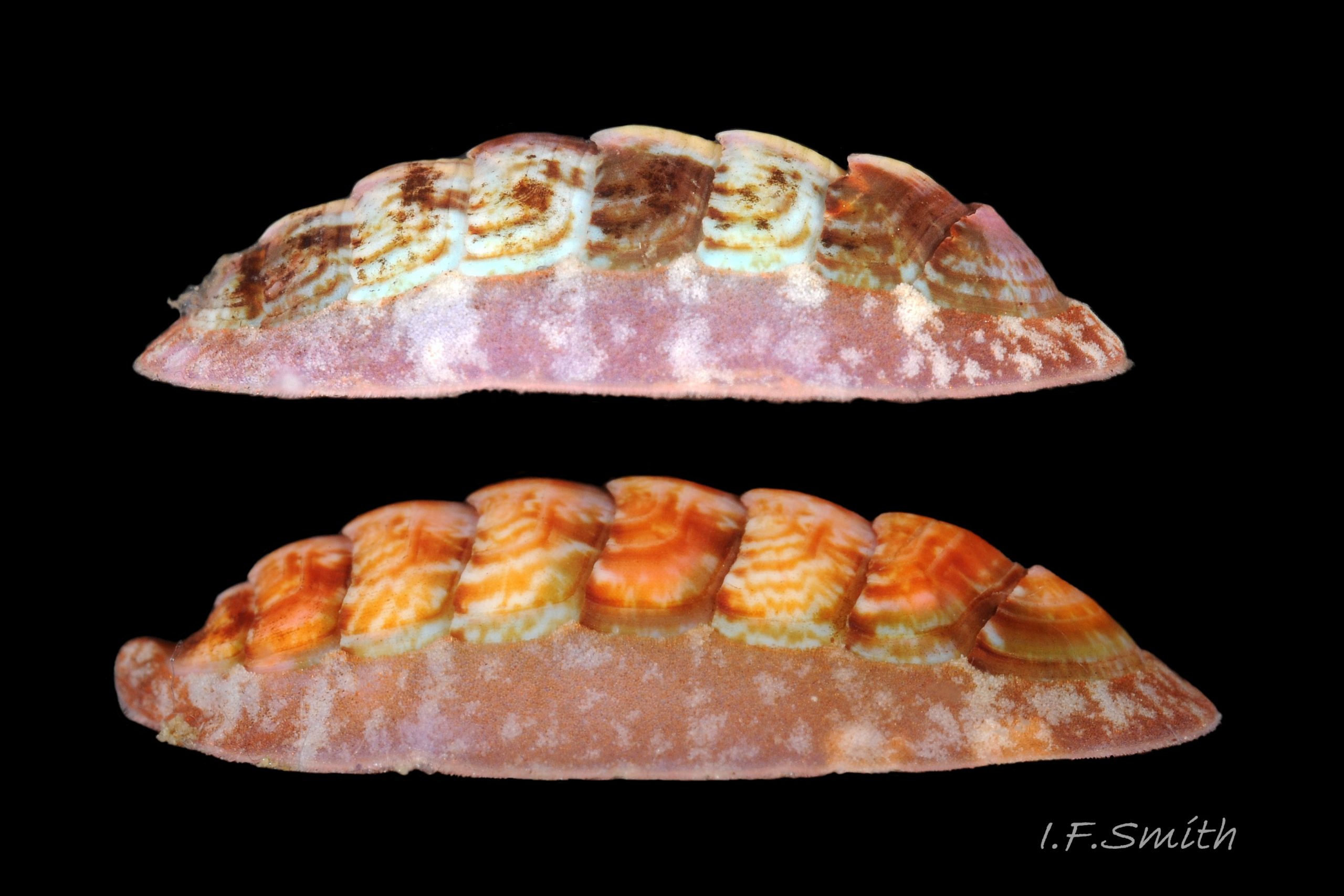
02 Boreochiton ruber
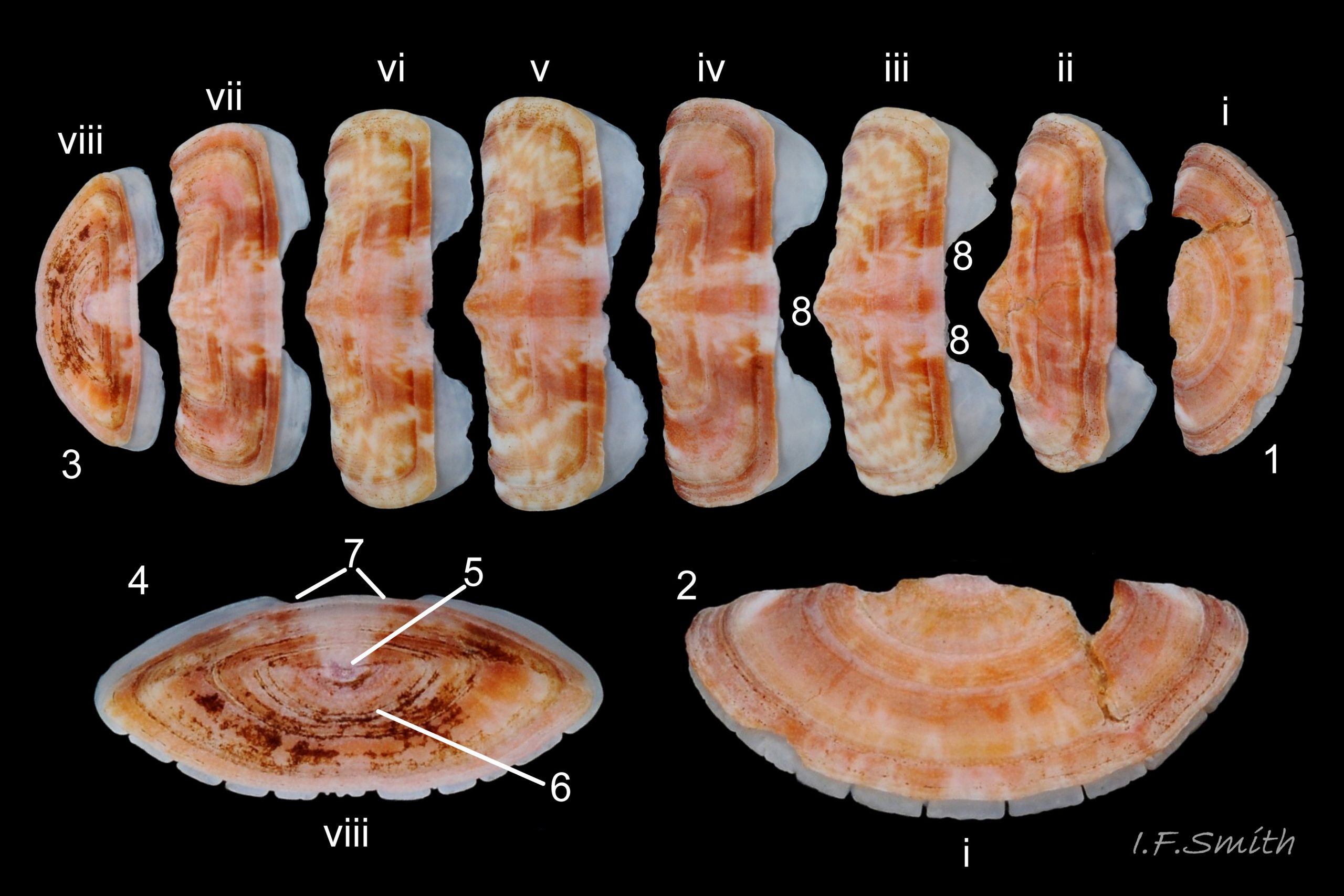
03 Boreochiton ruber
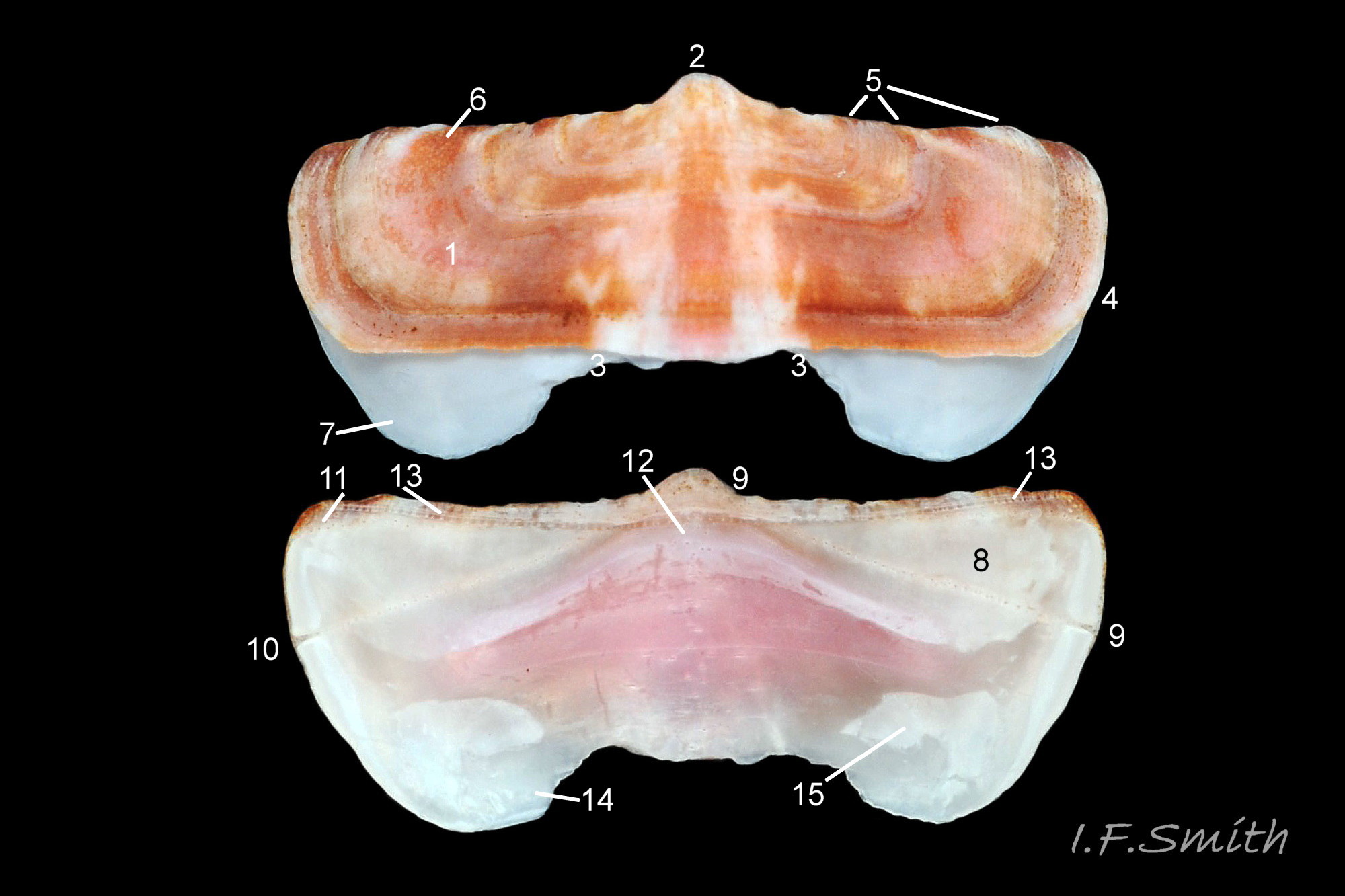
04 Boreochiton ruber
05 Boreochiton ruber
06 Boreochiton ruber
07 Boreochiton ruber

08 Boreochiton ruber
09 Boreochiton ruber
10 Boreochiton ruber
11 Boreochiton ruber
12 Boreochiton ruber
13 Boreochiton ruber
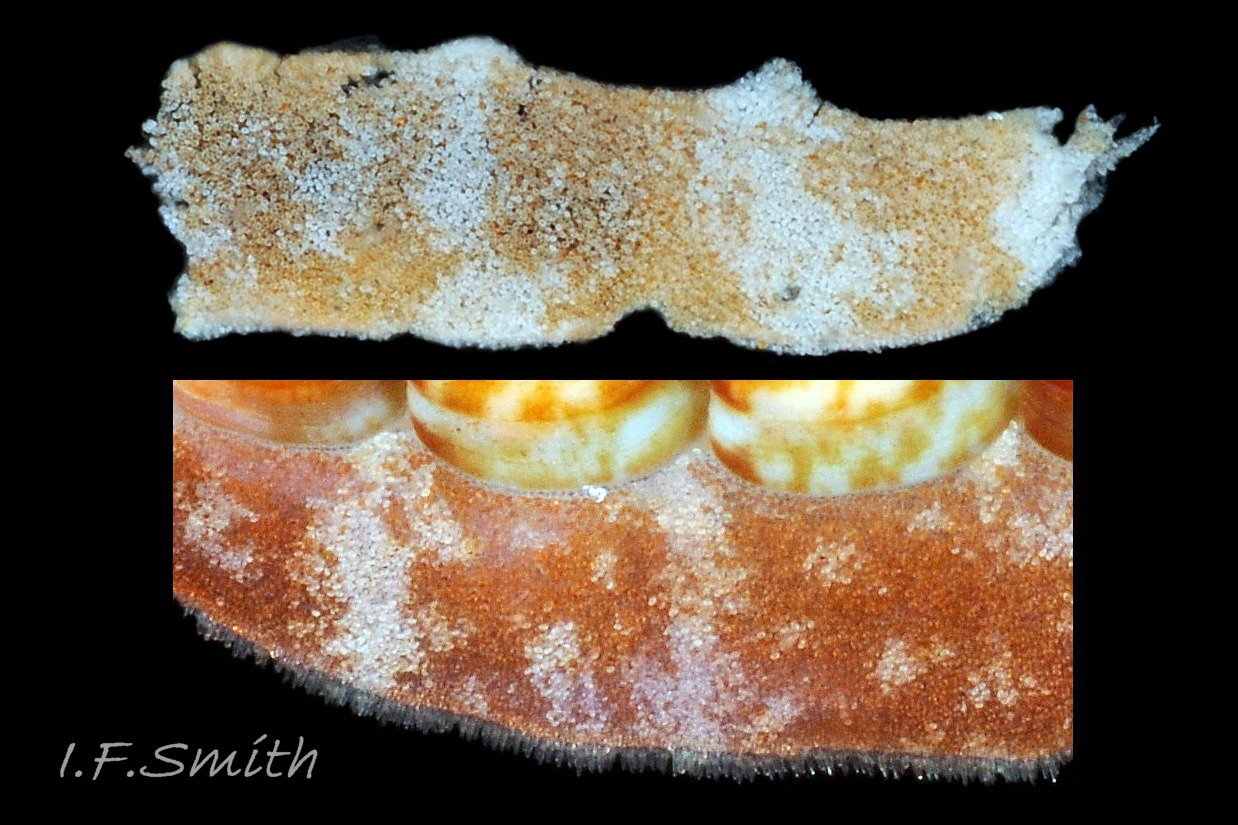
14 Boreochiton ruber
15 Boreochiton ruber
16 Boreochiton ruber
17 Boreochiton ruber COMPARISON Tonicella marmorea
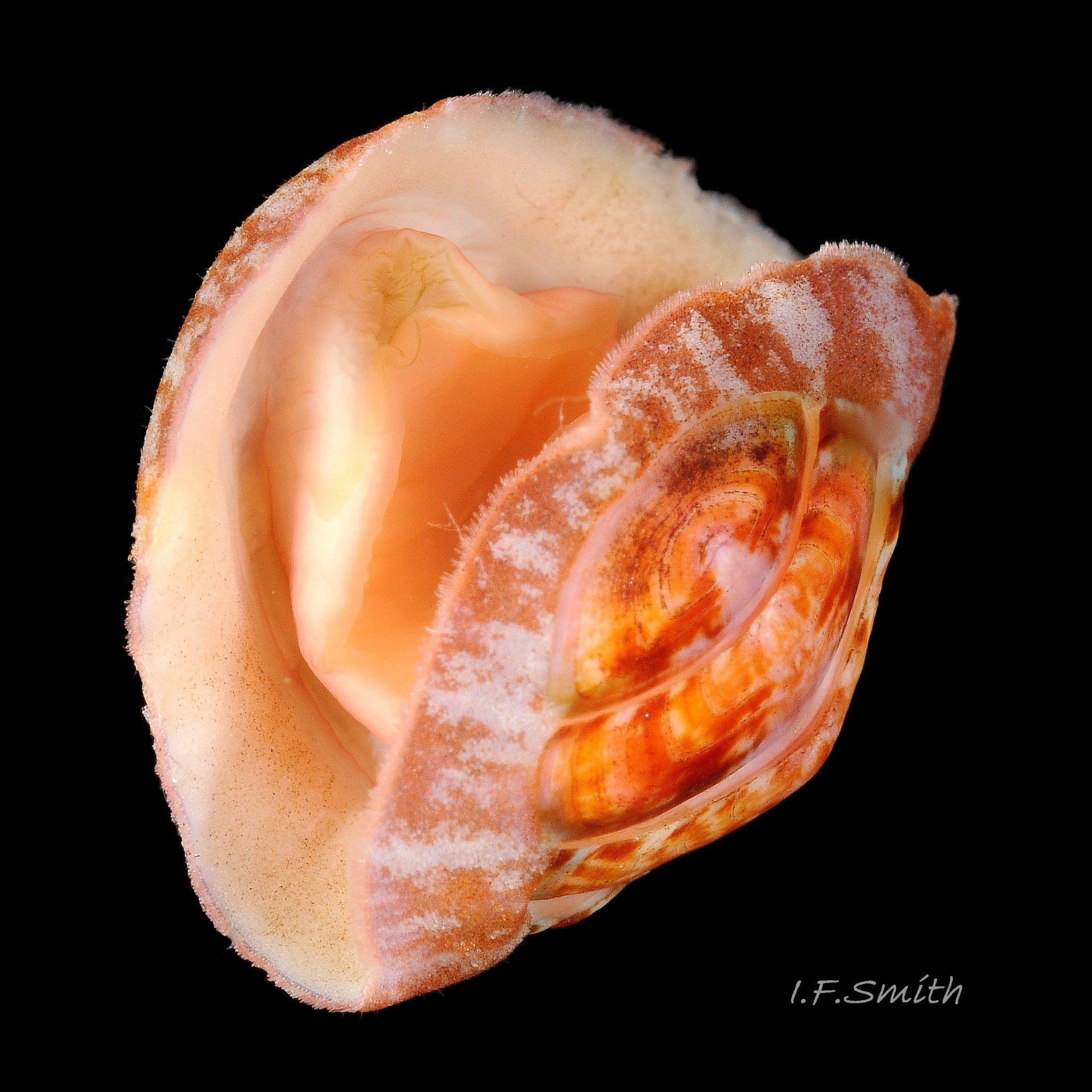
18 Boreochiton ruber

19 Boreochiton ruber

20 Boreochiton ruber COMPARISON image of Tonicella marmorea on left, chiton length 29mm, leg. S. Taylor & D. McKay, Lewis 2018. Boreochiton ruber on right, at same scale, chiton length 14.5mm, leg. P. Lightfoot, N. Yorkshire, 2014.

21 Boreochiton ruber COMPARISON L. cinerea

22 Boreochiton ruber COMPARISON Lepidochitona cinerea

23 Boreochiton ruber COMPARISON Lepidochitona cinerea
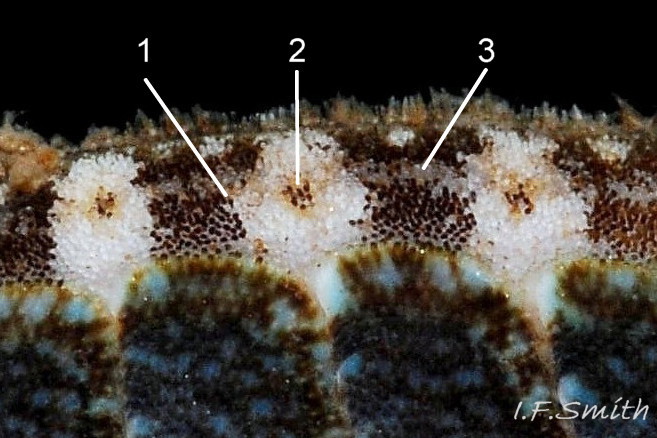
24 Boreochiton ruber COMPARISON Lepidochitona cinerea
Boreochiton ruber (Linnaeus, 1767) is currently (2019) accepted name.
Current taxonomy: World Register of Marine Species (WoRMS)
www.marinespecies.org/aphia.php?p=taxdetails&id=140152
Account was compiled using the now unaccepted name
Tonicella rubra (Linnaeus, 1767).
A pdf of this and other species accounts can be downloaded from www.researchgate.net/profile/Ian_Smith19/research
Authors: Paula Lightfoot and Ian F. Smith.
Synonyms:Chiton ruber Linnaeus, 1776; Chiton ruber var. oblonga Jeffreys, 1865; Tonicella rubra (Linnaeus, 1767).
Vernacular: Northern red chiton; Lleuen goch y graig (Welsh); Chiton rouge du nord (French); Rote Käferschnecke (German); Rødleddsnegl (Norwegian); Rød skallus (Danish);
GLOSSARY BELOW This account uses the standardised terminology for chitons proposed by Schwabe (2010). Some of Jones & Baxter (1987) alternatives are indicated in the glossary as a.k.a..
Shell Description
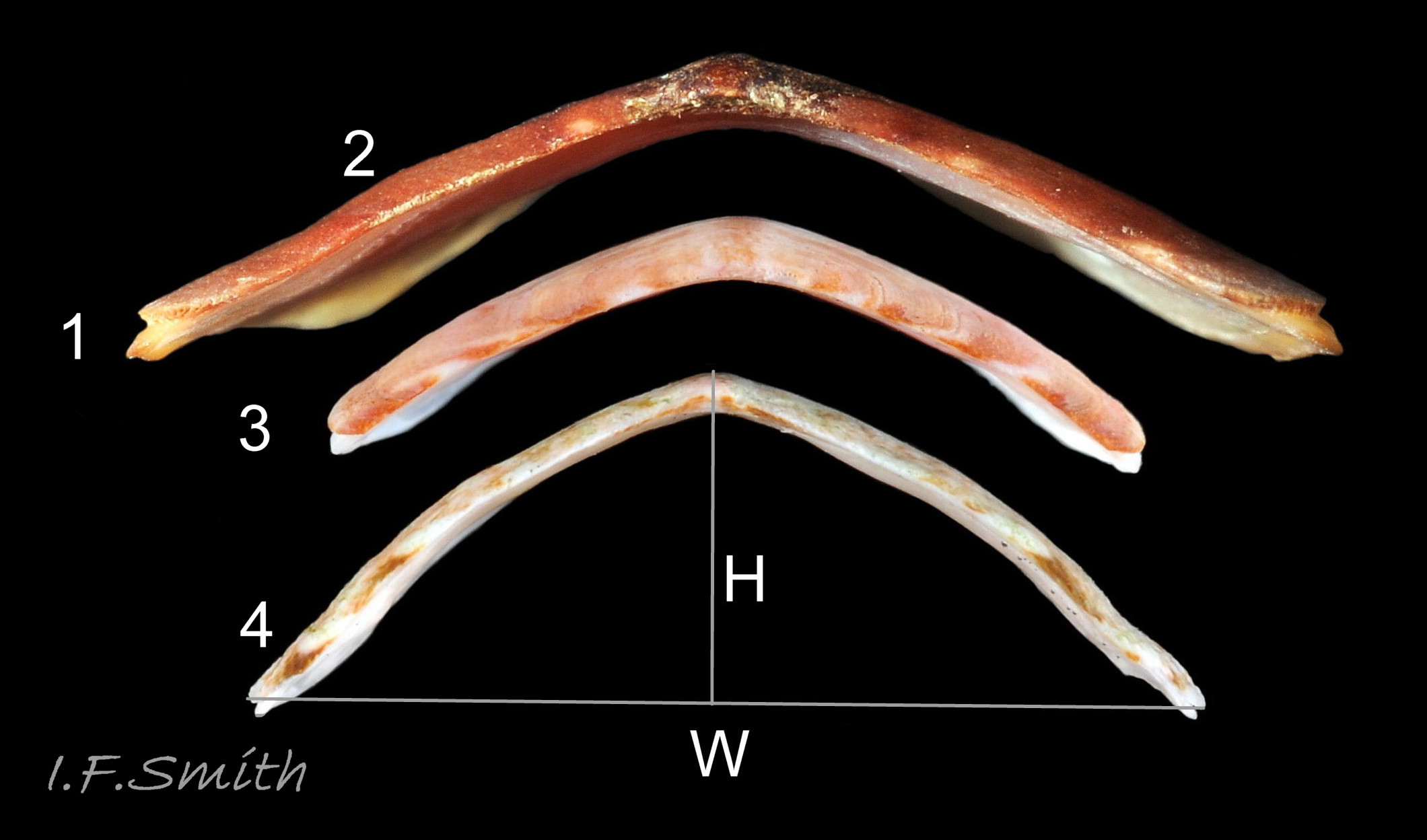
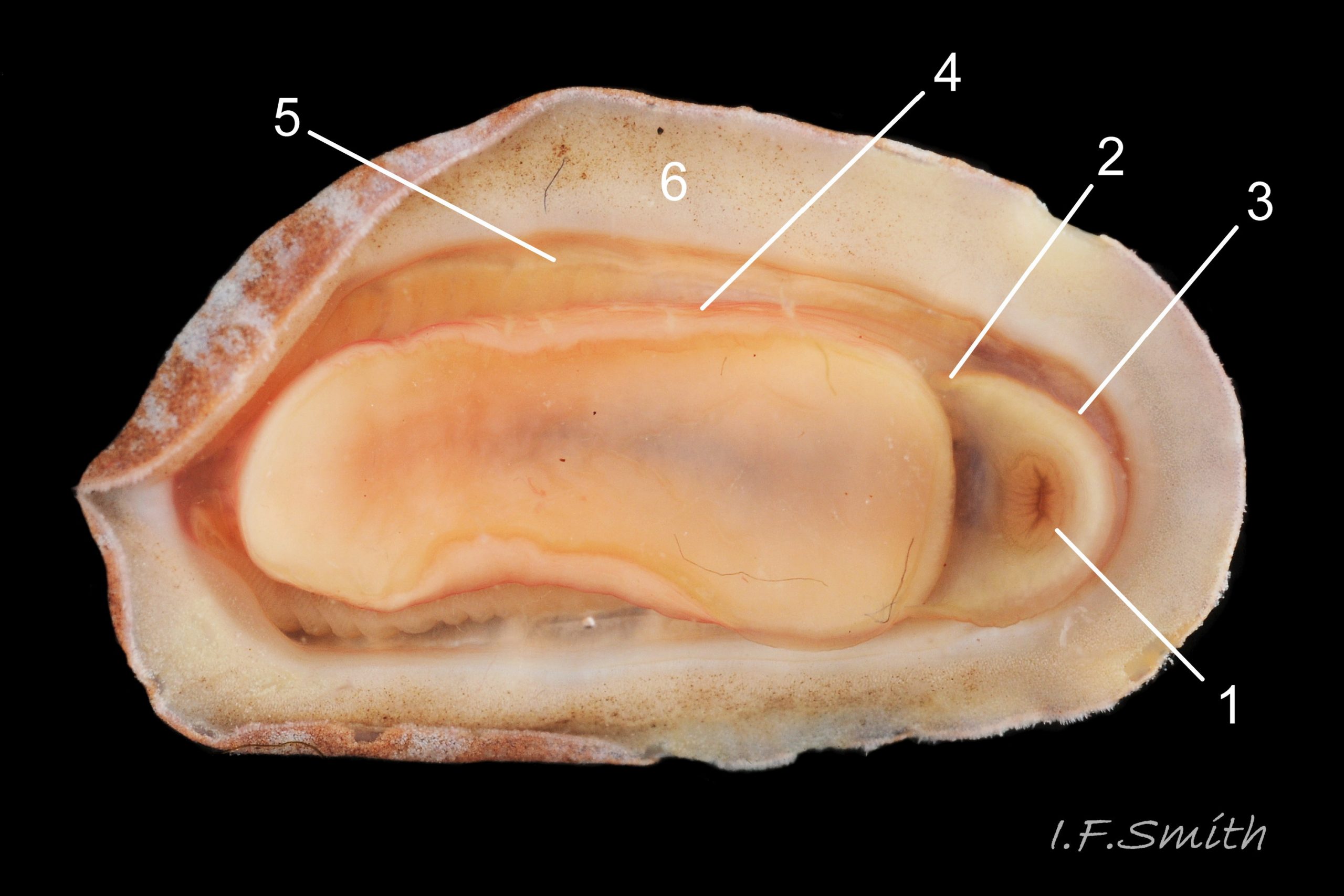
Usually 10 – 15 mm long and 6 – 9mm wide, occasionally 21mm long. Dorso-ventrally flattened, bilaterally symmetrical. In dorsal view, outline elliptical; width about 60% of length. 20-30% of animal width occupied by girdle (Jones & Baxter) 01 Boreochiton ruber, but in lateral view, because of inclined side slope of shell and perspective, girdle appears to take up about 50% of animal’s width 02 Boreochiton ruber .
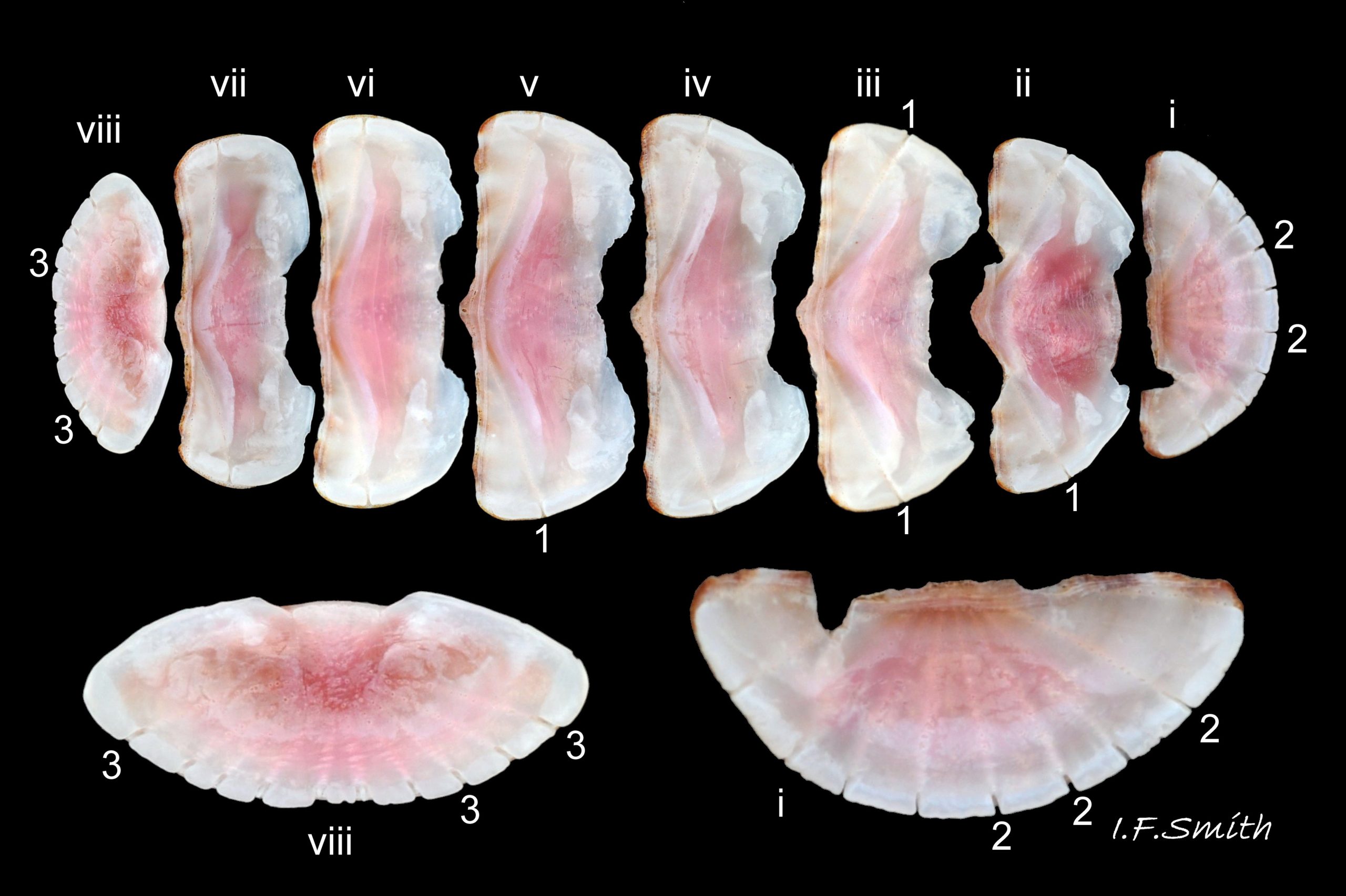
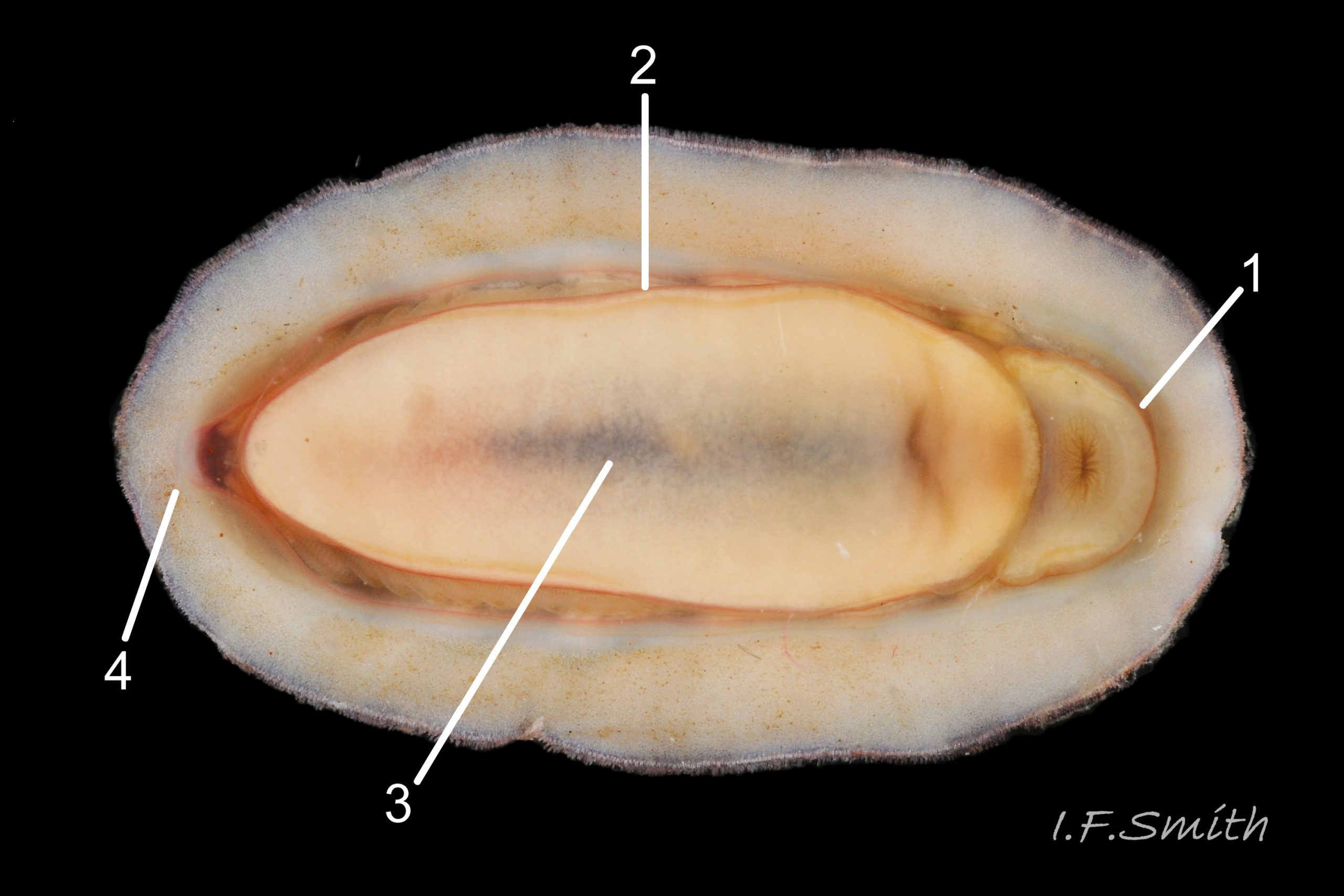
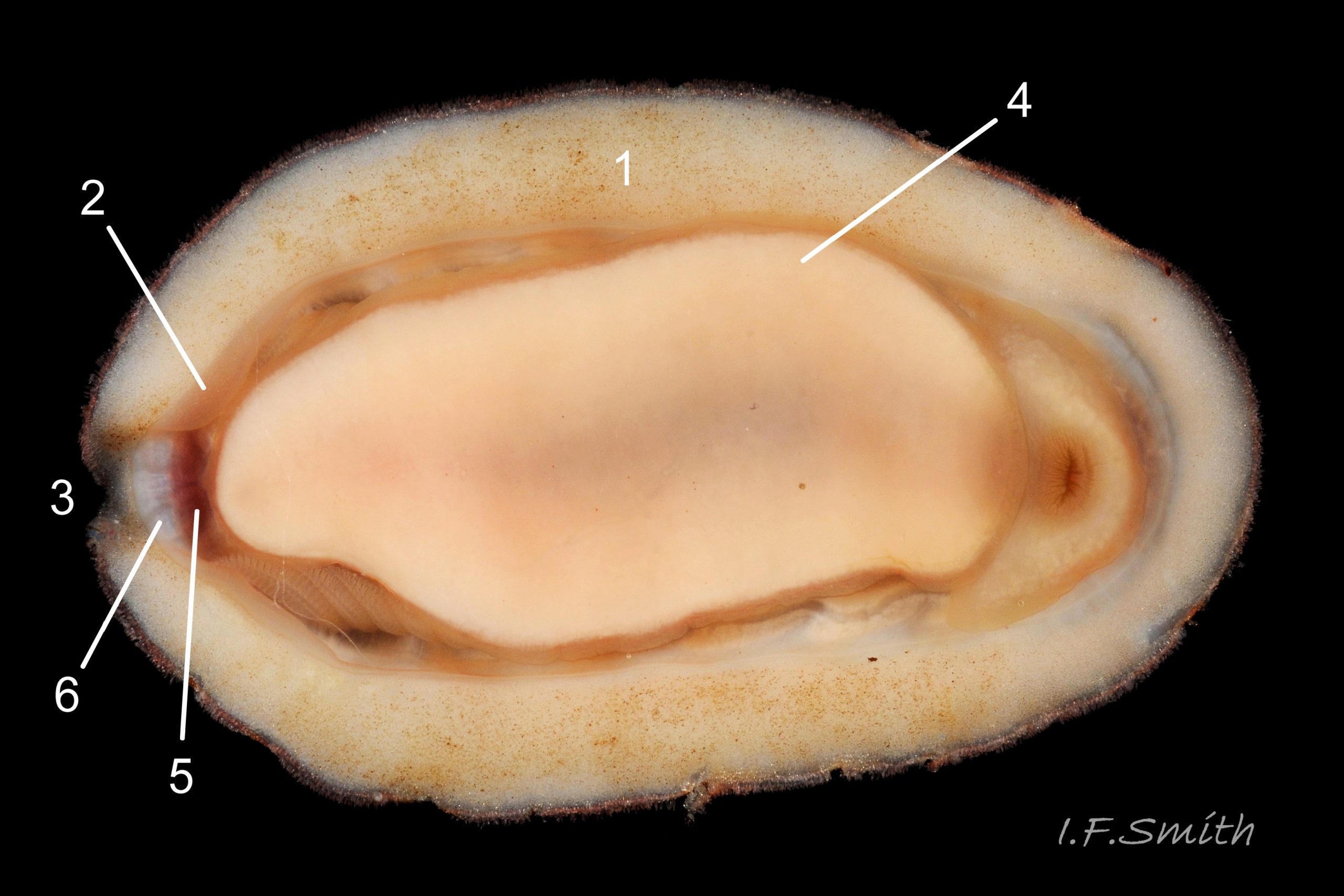
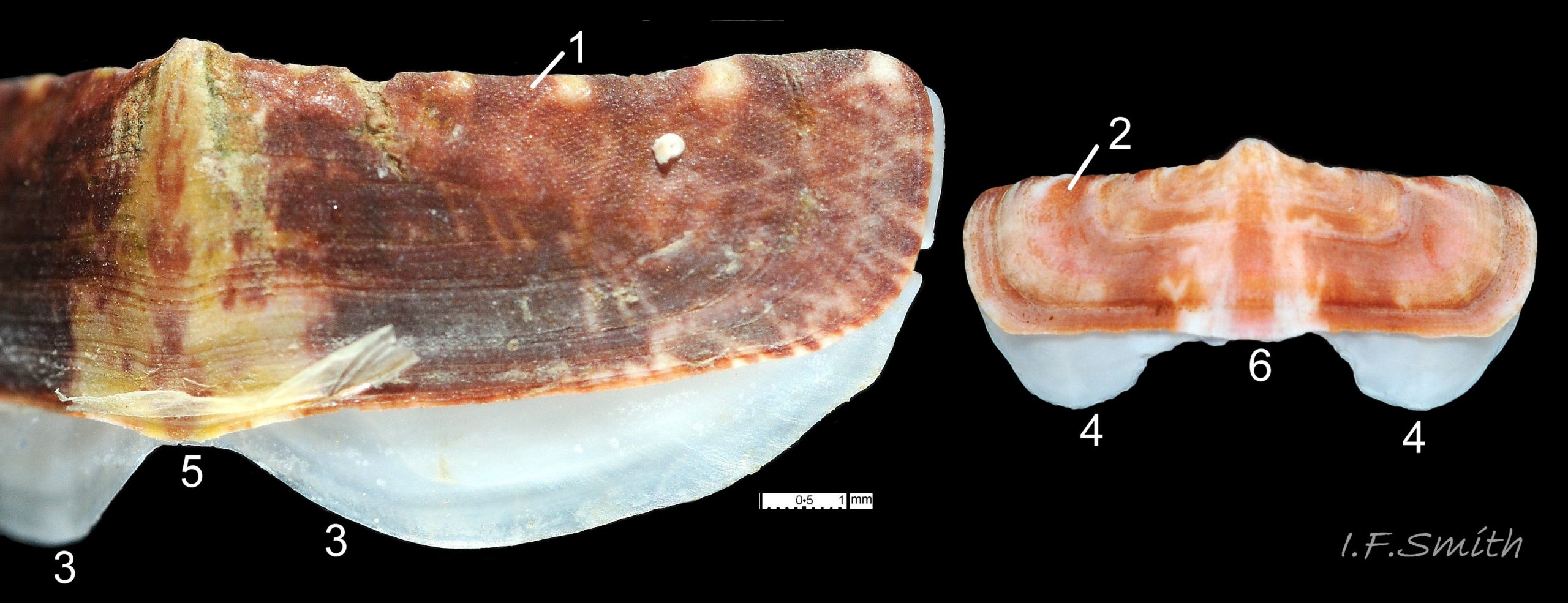
Eight overlapping valves 03 Boreochiton ruber principally made of 1) tegmentum: outer coloured layer of aragonite permeated and weakened by canals, and 2) articulamentum: thick inner layer of white and (on jugal tract) pink aragonite, also 3) properiostracum: outermost proteinaceous layer containing colour and 4) myostracum: thin discontinuous innermost layer (only visible in section examined under microscope) 04 Boreochiton ruber . Head valve (i) semicircular, with wide V posterior edge when viewed in situ on living specimen 01 Boreochiton ruber. Intermediate valves (ii –vii) rectangular, slightly keeled, with low smoothly-convex to straight side slopes when viewed from posterior 05 Boreochiton ruber ; noticeably lower arch-profile than Lepidochitona cinerea, but similar height to Tonicella marmorea. Less than semicircular tail valve (viii) has antemedian mucro with almost straight, steep postmucronal slope; antemucronal area resembles jugal area of intermediate valves 03 Boreochiton ruber . Distinct beak on each of intermediate valves (ii –vii) visible in lateral view 02 Boreochiton ruber , and mucro on tail valve viii resembles beak.
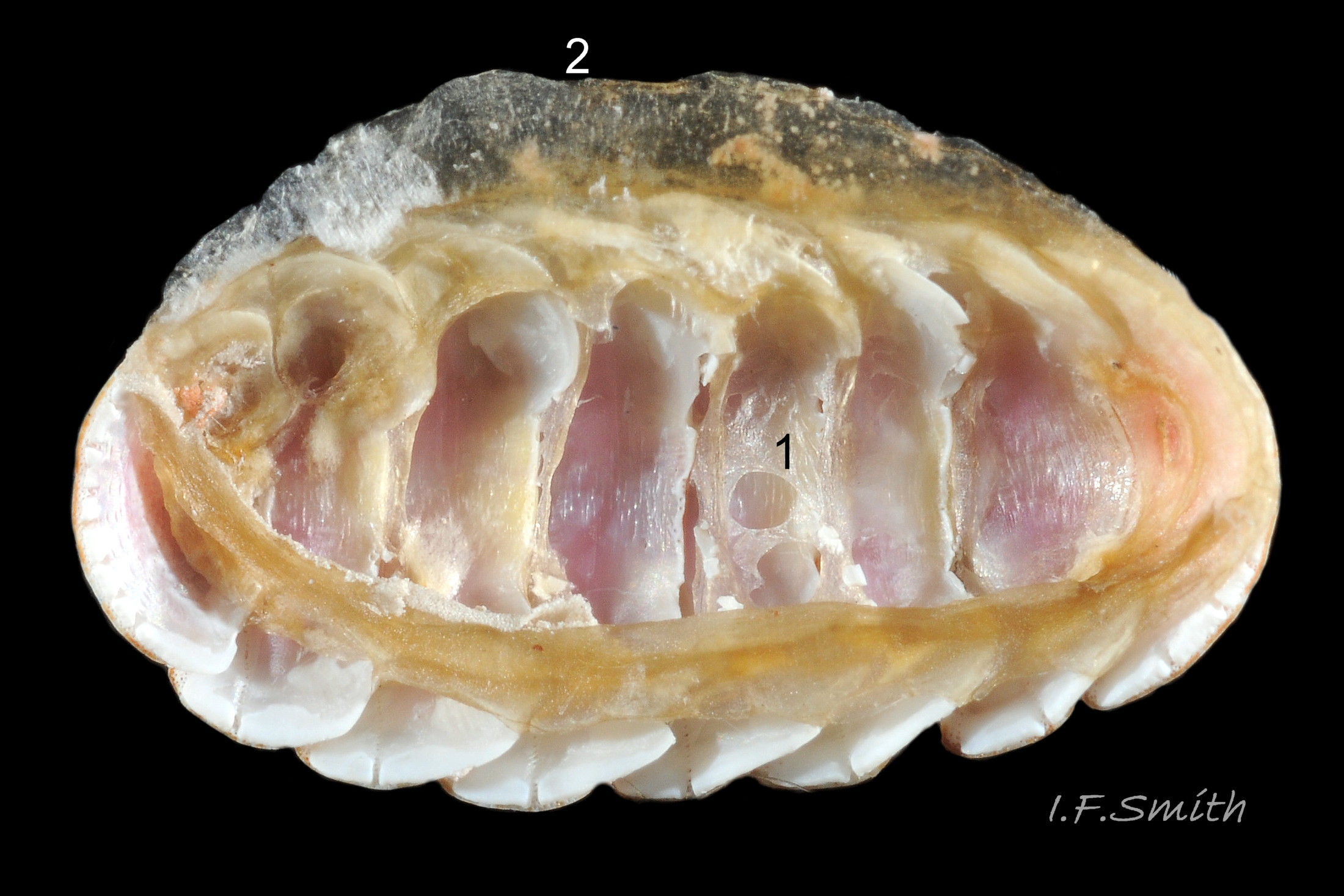
Canals permeate tegmentum and terminate on its dorsal surface as minute stipple of caps on pores (magnification over X30), and as open pores on margins of tegmentum; some canals penetrate the articulamentum to form holes on its ventral surface, especially near the posterior edge and in the slit ray and jugal tract 04 Boreochiton ruber .
Insertion plates of valves ii – vii embedded in girdle have a single slit; head valve (i) has about eight slits and tail valve (viii) has about nine or ten slits 06 Boreochiton ruber . Typical slit formula 8/1/9 or10, but can vary. Weakly developed slit rays run from slits to posterior edge of valves i – vii 04 Boreochiton ruber and from slits to mucro on valve viii 06 Boreochiton ruber .
Valves ii – viii each have two apophyses on anterior edge; those on valves iii –v are markedly prominent and rounded, almost semicircular. Tail valve (viii) has short (i.e. antero-posterior distance), wide, near-trapezoidal apophyses. 04 Boreochiton ruber . Apophyses extend under next valve forwards.
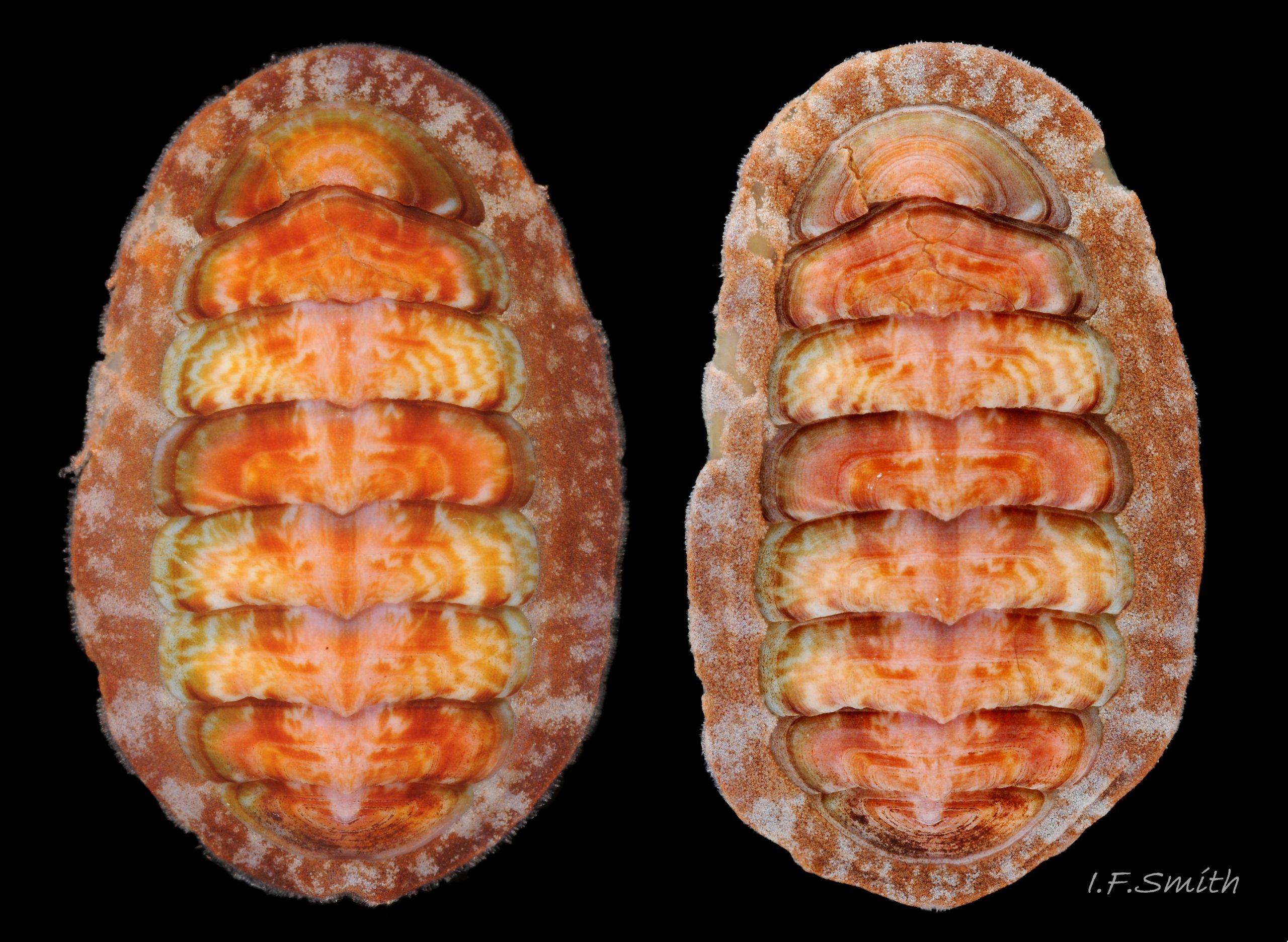
Dorsal surface of valves smooth apart from near-rectangular, distinct, growth lines (faint stippling of caps on canals may be visible at over X30 magnification); linear colouring differentiates jugal area, but lateral and pleural areas very weakly defined as separating diagonal ridge undeveloped 04 Boreochiton ruber . Shell growth is outwards from posterior margin of all valves, except concentrically outwards from mucro on tail valve (viii) 03 Boreochiton ruber . Properiostracum quite thick, has consistency like collagen, may become loose and easily rubbed off if animal loses condition in captivity. Dorsal shell-colour results partly from pigment in properiostracum; brightest on live specimens in good condition 07 Boreochiton ruber , vibrancy lost immediately upon death, or even before if in poor condition or injured 03 Boreochiton ruber & 08 Boreochiton ruber . When alive, principal colours of shell and girdle lie in range of brown, brick-red, orange, pink and purple-pink in various combinations arranged in streaks and blotches, with intervening areas of cream or greenish-cream 02 Boreochiton ruber & 09 Boreochiton ruber chemical stability of pigments varies; reds persist after death, while greens soon fade.
Body Description
Head and foot never protrude into view naturally on live animal, can only be seen if animal removed from substrate, or placed on glass.
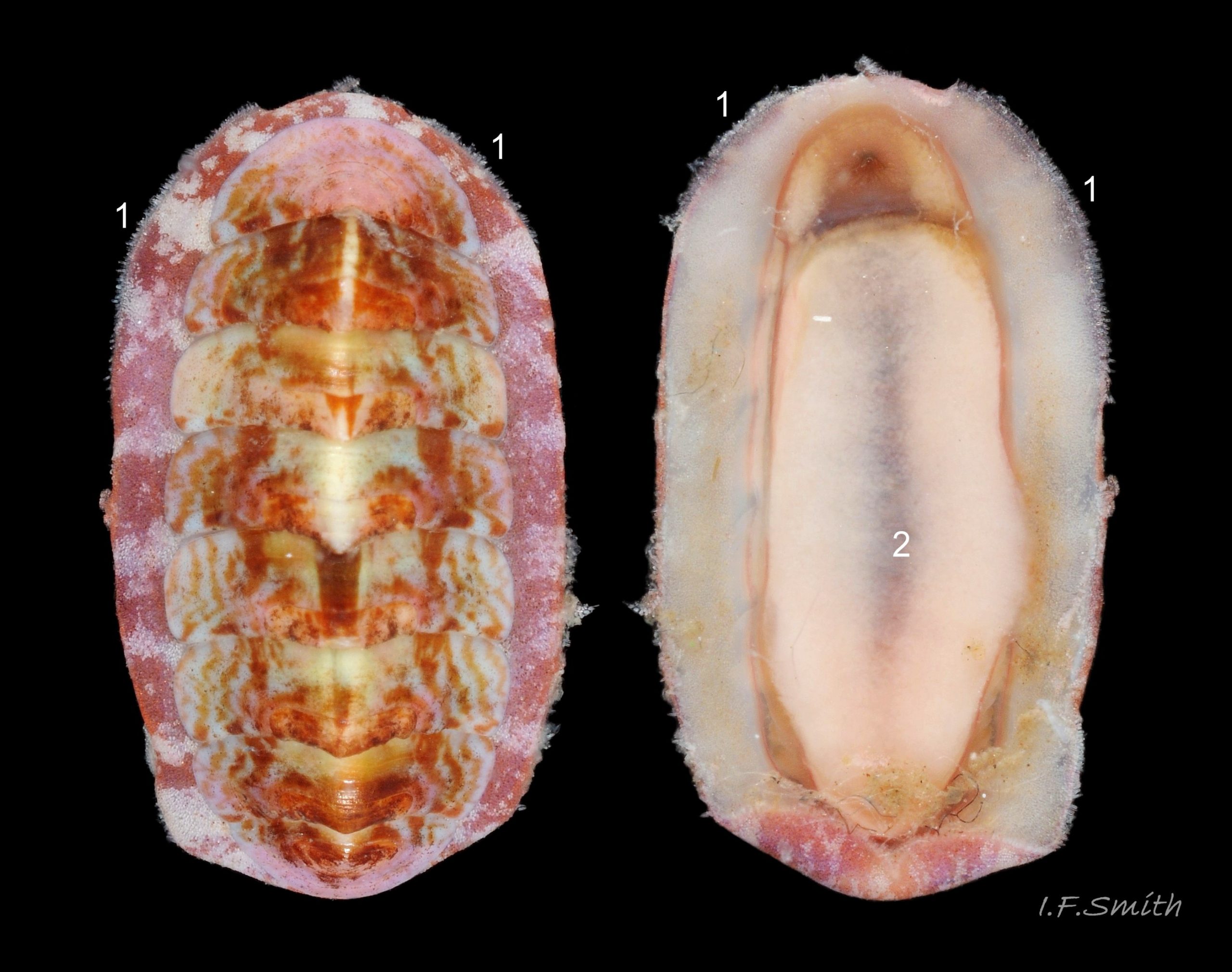
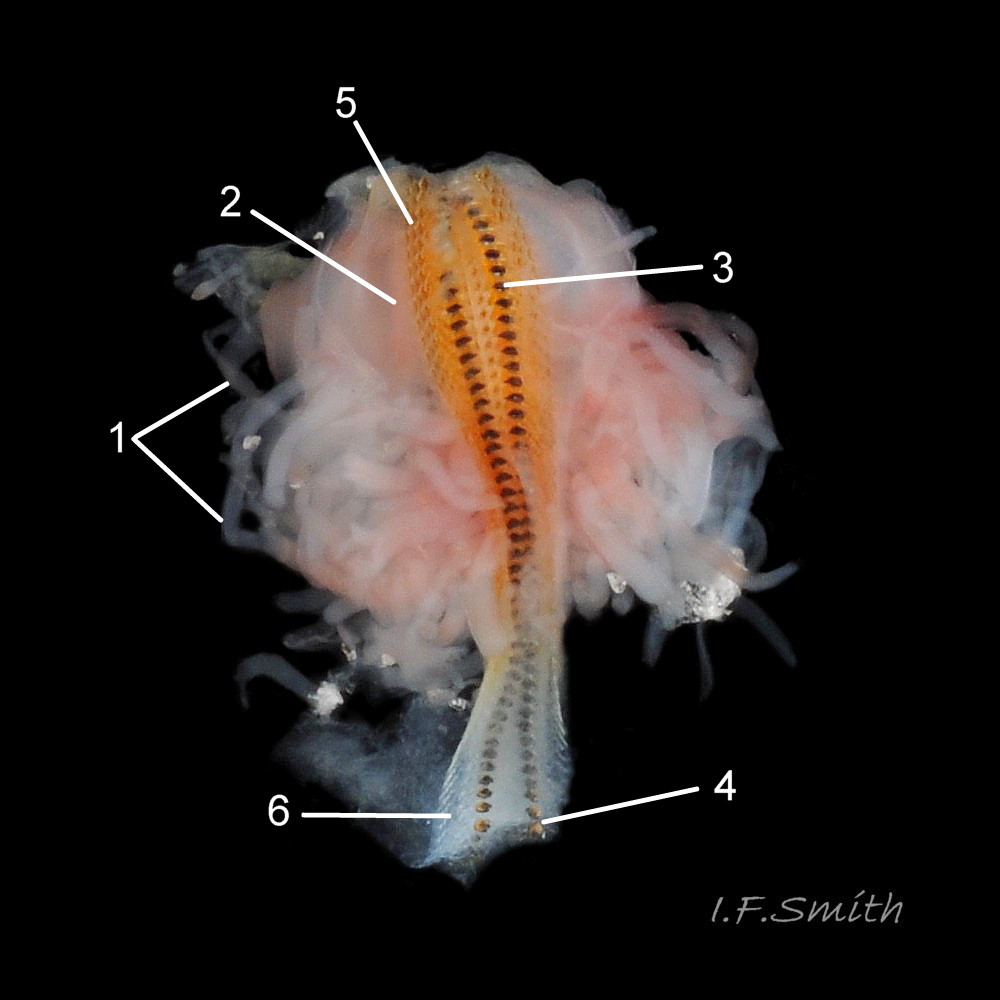
Head shaped as crescent moon with concave edge fitting round anterior of foot and extending laterally as mouth lappets 10 Boreochiton ruber . Much of head occupied by large transverse slit-mouth with wrinkled lips; no eyes or sensory tentacles. Pink 10 Boreochiton ruber or brownish-pink 11 Boreochiton ruber border around head when viewed ventrally 12 Boreochiton ruber . As on chitons generally, broad radula with extremely strong hard teeth of chitin mineralized with magnetite in rows of seventeen. Teeth pale when formed at rear of radula, darken and harden with magnetite as moved forwards. In each row a pair of major lateral teeth, the principal scrapers, are the largest and darkest 16 Boreochiton ruber .
Aesthetes (sensory tissue) fill canals that permeate the tegmentum and parts of articulamentum; terminate on fine stipple (may be visible at over 30X magnification 04 Boreochiton ruber ) as sense organs on dorsal surface of valves.
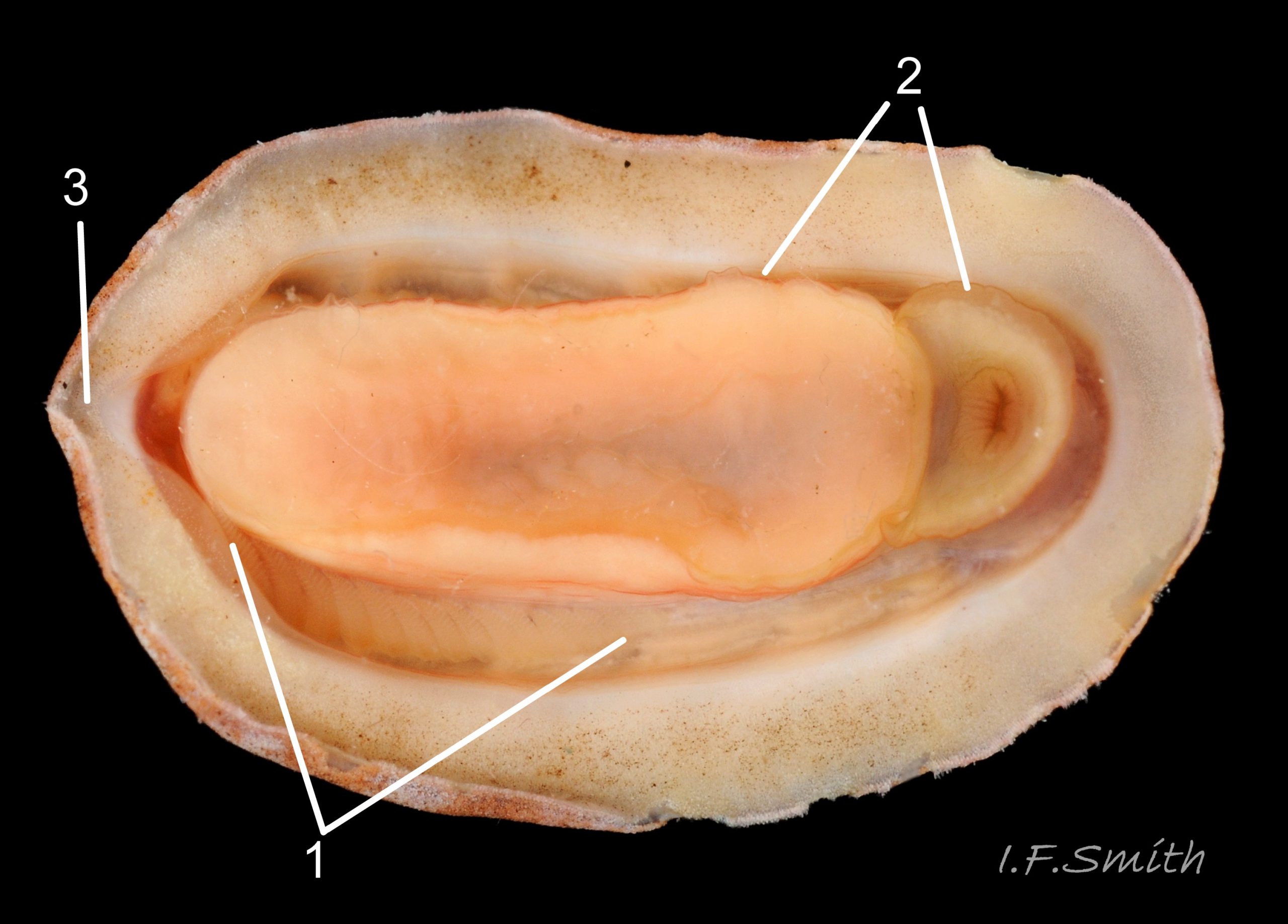
Immediately below the valves, mantle is tough thin translucent epidermis 13 Boreochiton ruber , but greatly thickened where reflected around periphery to form a fleshy girdle into which ends of valves deeply embedded 14 Boreochiton ruber . Girdle has cream or yellowish-white flesh with tough transparent yellowish epidermis that survives when flesh rotted away 13 Boreochiton ruber . Ventral surface of girdle (hyponotum) paved with elongate oval, strongly ribbed, colourless scales (20X16μm to 44×18μm), often some stained brownish 15 Boreochiton ruber . Dorsal surface of girdle (perinotum) has densely-packed, elongate (50 – 60μm), club-shaped granules with packed touching heads giving a mealy 14 Boreochiton ruber . Perinotum granules coloured similarly to valves with often-fragmentary, pale, narrow bands roughly in line with shell sutures 02 Boreochiton ruber . Edge of girdle fringed by straight, abruptly pointed, longitudinally finely striate spicules (124 X 22μm) when fresh and unworn, often tinted as girdle 09 Boreochiton ruber . Hyponotum can be flexed outwards to form channel at posterior for release of faecal pellets, ova or sperm 11 Boreochiton ruber , 12 Boreochiton ruber , 15 Boreochiton ruber and flic.kr/p/n6WTou (B. Forrest, related sp. T. marmorea)
Open narrow mantle-cavity runs around whole animal; contains about twelve small ctenidia (range 10 – 15) on each side in the posterior half only (merobranch arrangement) 12 Boreochiton ruber . Number of ctenidia increases with age. Between mantle-cavity and hyponotum the mantle-fold is unobtrusive 10 Boreochiton ruber except where widens near posterior into mantle-lappet 15 Boreochiton ruber and may partly conceal ctenidia. Anus opens into mantle-cavity at posterior by channel formed by deflection/ depression of hyponotum 15 Boreochiton ruber . Nephridiopores and gonopores open laterally into posterior quarter of cavity. No penis as external fertilization. Foot, elongate ellipse, has cream or yellowish-white sole with dark viscera sometimes visible medially 11 Boreochiton ruber , but no medial dividing line. Foot has pink 10Tr 10 Boreochiton ruber or brownish-pink border 11 Boreochiton ruber around sole when viewed ventrally. Foot spreads widely, concealing pallial cavity when gripping substrate 15 Boreochiton ruber .
Key identification features
·Boreochiton ruber .
·At LWS and sublittorally.
·Dorsal surface of valves smooth apart from growth lines. (Faint stippling may be visible at X30 magnification.) 17 Boreochiton ruber.
·Apophyses, especially on valves iii –v, are markedly prominent and rounded, almost semicircular 17 Boreochiton ruber.
·Dorsal surface of girdle has densely-packed, elongate (50 – 60μm), club-shaped granules with packed touching heads giving a mealy appearance. 14 Boreochiton ruber .
·Girdle usually has alternating dark and pale transverse bands. Pale bands, often fragmentary, usually thinner than dark bands, and roughly in line with shell-sutures 02 Boreochiton ruber .
·Dorsal surface of valves ii – vii makes an arch with smoothly-curved or flattish sides when viewed in profile from posterior 05 Boreochiton ruber . Arch distinctly lower than that of Lepidochitona cinerea, but similar to that on T. marmorea.
·about 12 ctenidia each side (range 10-15), arrangement merobranch12 Boreochiton ruber .
Similar species
For distinguishing from small T. marmorea (length 21mm or less), underwater photographs may not be sufficient; counting of ctenidia, careful disarticulation of specimen and examination under magnification may be required.
·Tonicella marmorea (O. Fabricius, 1780).
·At LWS and sublittorally.
·Dorsal surface of valves has growth lines and, at X5 magnification, numerous distinct small granules 17 Boreochiton ruber. (Granules, not as large as on L. cinerea, may not be discernible in underwater images or on specimens before deposits and properiostracum removed.)
·apophyses on intermediate valves short (i.e. anterior-posterior), wide and gently curved 17 Boreochiton ruber. (Note, apophyses in 4 in Kaas & Belle appear to be worn/damaged to stronger curve. Fig.19 in Jones & Baxter shows un-damaged apophyses more accurately.)
·Dorsal surface of girdle has sparse minute granules(c.27μm long) that do not touch each other. (Usually escape notice in photographs.) 20 Boreochiton ruber
·Girdle variably coloured and patterned; deep purple-red to green and may have orange bands; some may resemble B. ruber 20 Boreochiton ruber.
·Dorsal surface of valves ii – vii flat or weakly convex upper slope, separated by hip from slightly concave lower slope 05 Lepidochitona cinerea Low arch, similar height/width ratio to B. ruber.
·17 – 26 ctenidia each side, arrangement merobranch.
·Not found in Britain further south than North Wales or Northumberland. (Many erroneous records on mapping schemes as easily confused with T. rubra.)
Lepidochitona cinerea (Linnaeus, 1767).
1. Only chiton species likely to be found higher than MLW on British shores. Has diverse colour forms 21 Boreochiton ruber , some similar to B. ruber. 22 Boreochiton ruber. Usual maximum length 16mm, occasionally 28mm.
2. Dorsal surface of valves has numerous rounded, sometimes slightly elongated, granules visible at X5 magnification 23 Boreochiton ruber
3. White apophyses on intermediate valves are gently curved. Wide jugal sinus (gap between apophyses) on intermediate valves is about same width as one apophysis 17 Boreochiton ruber
Dorsal surface of girdle has densely packed rounded granules 23 Boreochiton ruber.
4. Girdle usually has alternating dark and light transverse lozenge-like bands of approximately equal size. Usually, dark bands have narrow waist, and light bands have a wide waist 24 Boreochiton ruber(sometimes colours interchanged). A thin paler greyish longitudinal line often runs across the waist of bands. Usually a dark central spot on pale bands and white spot somewhere on dark band. Markings can be partial, indistinct, or absent on pale specimens.
5. Valves moderately highly arched; valve 5 height about 32% of width; distinctly higher than that of B. ruber 05 Boreochiton ruber
6. Usually 16 – 19 gills each side, arrangement holobranch.
Habits and ecology
Lives on and under stones and old shells, including those on sandy seabed, and on rock around holdfasts of Laminaria at LWS and sublittorally to about 300 metres, commonest 10 – 40m. Shell articulates to conform closely to uneven rock surface; large foot and flat hyponotum grip surface firmly. When alarmed, can increase grip suctorially by raising part of hyponotum to form partial vacuum, and if displaced from substrate, can roll into a ball 18 Boreochiton ruber . Respiration: cilia on ctenidia and mantle create inhalent water-current entering pallial cavity wherever girdle is raised at anterior. Adjacent ctenidia have interlocking cilia so all work as a unit (as in many bivalves). Water current passes through ctenidia and then along cavity as exhalent current to posterior to exit through channel formed by deflection of hyponotum of girdle 15 Boreochiton ruber . Circulation: blood travels from head sinus through longitudinal sinuses to the foot, viscera and mantle, then through the ctenidia 12 Boreochiton ruber for aeration before passing into the adjacent heart in the dark-red dorso-ventrally flattened pericardium below valves vii & viii (occasionally exposed on live specimens 15 Boreochiton ruber ). From the heart the blood is pumped through the medial dorsal aorta to the head sinus, giving off some channels to the gonad and valve muscles on the way. In absence of eyes, or of tactile or chemoreceptor tentacles on head, senses environment through aesthetes exposed on surface of shell. Precise functions of aesthetes uncertain, but focusing lens and receptive retina found in aesthetes of some chiton species, though not on the related Tonicella marmorea (may also apply to T. rubra). Other proposed aesthete functions include chemoreception, mechanoreception, replenishment of properiostracum materials, and secretion of protective substances. Also sensory organs on girdle.
Feeds by scraping microalgae and associated organisms from the rock surface using its hard radula of chitin mineralized with magnetite 16 Boreochiton ruber . Though primitive in several ways, has elaborate digestive system with no trace of primitive rotating style in stomach. Long coiled intestine compacts faeces into oval pellets flic.kr/p/n6WTou (B. Forrest, related T. marmorea) . Water current in pallial cavity carries excreta from lateral nephridiopores to posterior, where faecal pellets from anus join the flow; all expelled at posterior through channel in hyponotum of girdle 15 Boreochiton ruber .
Travels by monotaxic retrograde compression waves on sole of foot.
Breeding: dioecious. Water current in pallial cavity carries sperm or ova from lateral gonopores to posterior and out through channel in raised girdle 15 Boreochiton ruber (images by B. Forrest of related T. marmorea : female releasing ova flic.kr/p/n6UtHY and male emitting sperm in gelatinous string from posterior flic.kr/p/n6WTou ). As fertilization is external, synchronised emission of sperm and ova needed to ensure success; trigger in many chiton species is moon-phase/ state of tides. Planktonic trochophore larvae hatch and metamorphose into small adult-form young without intervening veliger stage.
Distribution and status
Widespread but rarely abundant. Greenland, Iceland, N. Norway and White Sea to Brittany and New England (USA); most numerous in northern part of range. Not Baltic or southern North Sea. Also reported from N. Pacific, but some there have distinctly granulated valve surfaces in contrast to smooth (or finely pustulate at X30) surfaces on T. rubra in Atlantic, so some doubt about identity . GBIF map www.gbif.org/species/6487696 . All around Britain and Ireland except Liverpool Bay and east coast Flamborough to Kent, NBN UK map species.nbnatlas.org/species/NHMSYS0021185045
Acknowledgements
Thanks are due to Boyd Forrest for access to his images of spawning chitons. I am indebted to Dr Julia Sigwart, Director of the Marine Laboratory, Queens University, Belfast, for detailed advice with the text and interpretation of the images. Any errors or omissions are the responsibility of I.F.S.
Links and references
Botelho, A. 2013. Zoologger: mollusc grows hardest teeth in the world New Scientist 3rd October 2013 www.newscientist.com/article/dn24329-zoologger-mollusc-gr…
Fernandez, C.Z., Vendrasco, M.J. & Runnegar,B. 2007. Aesthete canal morphology in twelve species of chiton (Polyplacophora). The Veliger 49(2) 51 – 69 www.researchgate.net/publication/246548141_Aesthete_Canal…
Forbes, E. & Hanley S. 1849-53. A history of the British mollusca and their shells. vol. 2 (1853) London, van Voorst. (As Chiton ruber; Free pdf at archive.org/stream/historyofbritish02forbe#page/398/mode/2up Use slide at base of page to select pp.399-402.)
Fox, R. 2007. Invertebrate Anatomy On Line. Katharina tunicata lanwebs.lander.edu/faculty/rsfox/invertebrates/katharina….
Hayward, P.J. & Ryland, J.S. 1995. Handbook of the marine fauna of north-west Europe. Oxford, Oxford University Press.
Jardim, J.A. & Simone, L.R.L. 2010. Redescription of Hanleya brachyplax (Polyplacophora, Hanleyidae) from the south-southeastern Brazilian coast. Pap. Avulsos Zool. (São Paulo) 50 no.40
www.scielo.br/scielo.php?pid=S0031-10492010004000001&…
Jeffreys, J.G. 1862-69. British conchology. vol. 3 (1865). London, van Voorst. (As Chiton ruber; Free pdf at archive.org/stream/britishconcholog03jeffr#page/224/mode/2up . Use slide at base of page to select pp.224-226.)
Jones, A.M. & Baxter, J.M. 1987. Molluscs: Caudofoveata, Solenogastres, Polyplacophora and Scaphopoda London, Linnean Society, and Estuarine and Brackish-water Sciences Association.
Kaas, P. & Van Belle, R.A. 1985. Monograph of living chitons Vol 1. Viderup, Denmark. Brill. Preview at
books.google.co.uk/books?id=CKautf2EUPkC&pg=PA7&s…
Matthews, G. circa1954. The identification of British chitons. Papers for Students No.9. London, Conchological Society of Great Britain and Ireland.
Schwabe, E. 2010. Illustrated summary of chiton terminology. Spixiana 33(2):171-194.
Free pdf at verlag-pfeil.de/04biol/pdf/spix33_2_02.pdf
Glossary
μm – 0.001 mm
aesthetes – complex of canals and cavities filled with sensory tissue that permeate tegmentum and locally penetrate articulamentum. Open as sensory pores on dorsal surface of valves; probably compensate for lack of sensory structures on head. Some sense light; other sensory function(s) uncertain, but various authors have variously proposed chemoreception, mechanoreception, properiostracum replenishment and secretion of protective substances.
afferent – (adj. of vessel) carrying blood etc towards an organ.
a.k.a. – also known as.
antemedian – (syn. antemedial) situated to anterior of middle.
antemucronal area – area situated to anterior of mucro.
apophysis – (pl. apophyses) natural protruberance within a shell for attachment of muscles. On chitons, anterior extension of articulamentum which underlies preceding valve; on all valves except head valve (i).
aragonite – orthorhombic crystalline mineral-form of calcium carbonate www.minerals.net/mineral/aragonite.aspx . Less common on land than calcite, but, currently, the more frequent mineral-form in oceans and living mollusc shells.
articulamentum – inner shell-layer of chiton valves, usually hard, white, porcelaneous aragonite and often differently coloured in central part. (Partially underlain by inconspicuous myostracum layer.)
buccal mass – anterior of digestive system including an odontophore that supports anterior of radula, and a complex of muscles to operate them and other mouthparts. Usually red or pink.
cephalic – (adj.) of or on the head.
chemoreception – sensing of chemicals; “smell / taste”.
chitin – semitransparent flexible horny protein.
chitinous – (adj.) made of chitin.
cilia (pl.) –motile linear extensions of membrane used in locomotion, or to create water currents in feeding. (“cilium” singular).
coll. – in the collection of (named person or institution) (cf. legit).
ctenidium – comb-like molluscan gill; usually an axis with a row of filaments either side.
dioecious – having separate male and female individuals, not hermaphrodite.
dorso-ventrally flattened – as if pressed flat from above.
efferent – (adj. of vessel) carrying blood etc away from an organ.
emersed – above water surface, not covered in water.
epithelium – tissue forming outer layer of body surface, “skin”.
geotactic – (adj.) of species that moves towards pull of gravity (positively geotactic) or away from it (negatively geotactic). (synonym: gravitaxic.)
girdle – (on chiton) peripheral band of thickened, reflexed mantle that encloses ends of valves.
gonopore – genital opening through which eggs or sperm are released.
holobranch – (of chitons) ctenidia in mantle-cavity extend full length of foot.
hyponotum – ventral surface of chiton’s girdle.
immersed – covered in water.
insertion plate – (on most chitons) extension of articulamentum on lateral margin of intermediate valves, anterior margin of head valve and posterior margin of tail valve. Inserts into, and anchors valve to, the girdle muscle block.
intermediate valve – (of chiton) any valve (ii – vii), except head valve (i) and tail valve (viii).
interspecific – (adj.) existing or arising between different species.
intraspecific – (adj.) occurring within a single species or involving members of one species.
jugal area – triangular middle section of central area of intermediate valves, with apex pointing to posterior; discernible when defined by differences of colour and/or sculpture (dorsal surface).
jugal tract – triangular middle section of central area of intermediate valves, with apex pointing to posterior; discernible when defined by densely arranged aesthete pores (ventral surface).
jugum – triangular middle section of central area of intermediate valves. (See jugal area and jugal tract.)
lateral area – (on intermediate valve of chiton) triangular area with its base along lateral edge of valve and its apex near the centre of the posterior edge. a.k.a. lateral triangle.
legit – (abbreviation; leg. or lgt.) collected/ found by (cf. coll.)
LWS – low water spring tide, two periods of a few days each month when tide falls lowest.
mantle – sheet of tissue that secretes the shell, surrounds the viscera and forms a cavity for the gill in most marine molluscs.
mantle-cavity – (on chitons) a.k.a. pallial groove; narrow groove around whole foot and head, roofed by mantle and containing ctenidia, nephridiopores and gonopores.
magnetite – mineral of iron oxide, one of hardest materials made by any living organism.
mechanoreception – sensing of touch, sound, pressure change and/or posture.
microvilli (pl.) – microscopic cellular membrane protrusions that increase the surface area of cells. Possible functions include absorption, secretion, cellular adhesion and mechanotransduction. (singular, microvillus).
MLW – mean low water mark.
monotaxic – (of locomotion waves on foot) single series of waves; each wave extends across complete width of foot.
merobranch – (of chitons) gills in pallial cavity only in posterior two-thirds of animal.
motile – (adj.) able to move spontaneously; applicable to whole animals or to parts of them such as cells, gametes or cilia.
mucro – projection on tail valve (viii) of chiton demarking posterior from rest of valve. Varies in prominence and position.
myostracum – thin, inconspicuous, discontinuous, innermost layer of chiton shell.
nephridium – tubular glandular excretory/ osmoregulatory organ. a.k.a. kidney.
nephridiopore –opening of nephridium for excretion. a.k.a. nephropore, or renal pore.
odontophore – firm, approximately ellipsoid, structure of cartilage supporting radula. Protruded like a tongue to operate radula.
osmoregulation – regulation of osmotic pressure to keep organism’s fluids from becoming too diluted or concentrated.
pericardium – membranous sac containing heart and start of aorta.
perinotum – dorsal surface of chiton’s girdle.
pleural area – (on intermediate valve of chiton) triangular area with its base along anterior edge of valve and its apex near the centre of the posterior edge. a.k.a. median triangle.
postmucronal – situated to posterior of mucro on tail valve.
properiostracum – (on chitons) proteinaceous material covering the shell. Different composition from periostracum of most other molluscs.
phototactic – (adj.) of species that moves towards light (positively phototactic) or away from it (negatively phototactic).
plankton – animals and plants that drift in pelagic zone (main body of water).
radula – ribbon of chitin bearing chitinous teeth that is extruded on a tongue-like odontophore of cartilage to rasp food. On chitons, teeth are usually impregnated with magnetite, a hard magnetic mineral of iron.
rec. – recorded by (person who submitted record, may be different from leg. and coll. persons/institution).
retrograde – (of locomotion waves on foot) waves travel from anterior to posterior.
side slope – (on chiton) shape in profile view (from posterior or anterior) of lateral areas of intermediate valves; may be straight, convex, concave or a combination of these.
sinus – dilated channel or receptacle containing blood etc.
slit ray -– row of canal pores running diagonally from lateral slit to posterior edge on ventral surface of chiton valve. a.k.a. notch ray.
suture – (of chiton) line where two valves meet.
taxis – directional locomotary response to external stimuli such as light, gravity, temperature or chemicals.
tegmentum – outer shell-layer of chiton valves, usually porous and relatively soft. (Covered by properiostracum when live.)
trochophore – spherical or pear-shaped larvae that move with aid of girdle of cilia. Stage preceding veliger, passed within gastropod egg in most spp. but free in plankton for limpets, Trochidae, Tricolia pullus and (with no veligers) chitons.
major lateral teeth– largest two teeth in each row of seventeen on chiton radulae; principal scraping blades. Often darkened more than other teeth by high magnetite content.
veliger – shelled larva of marine gastropod or bivalve mollusc which swims by beating cilia of a velum (bilobed flap).