Click image to enlarge with full caption
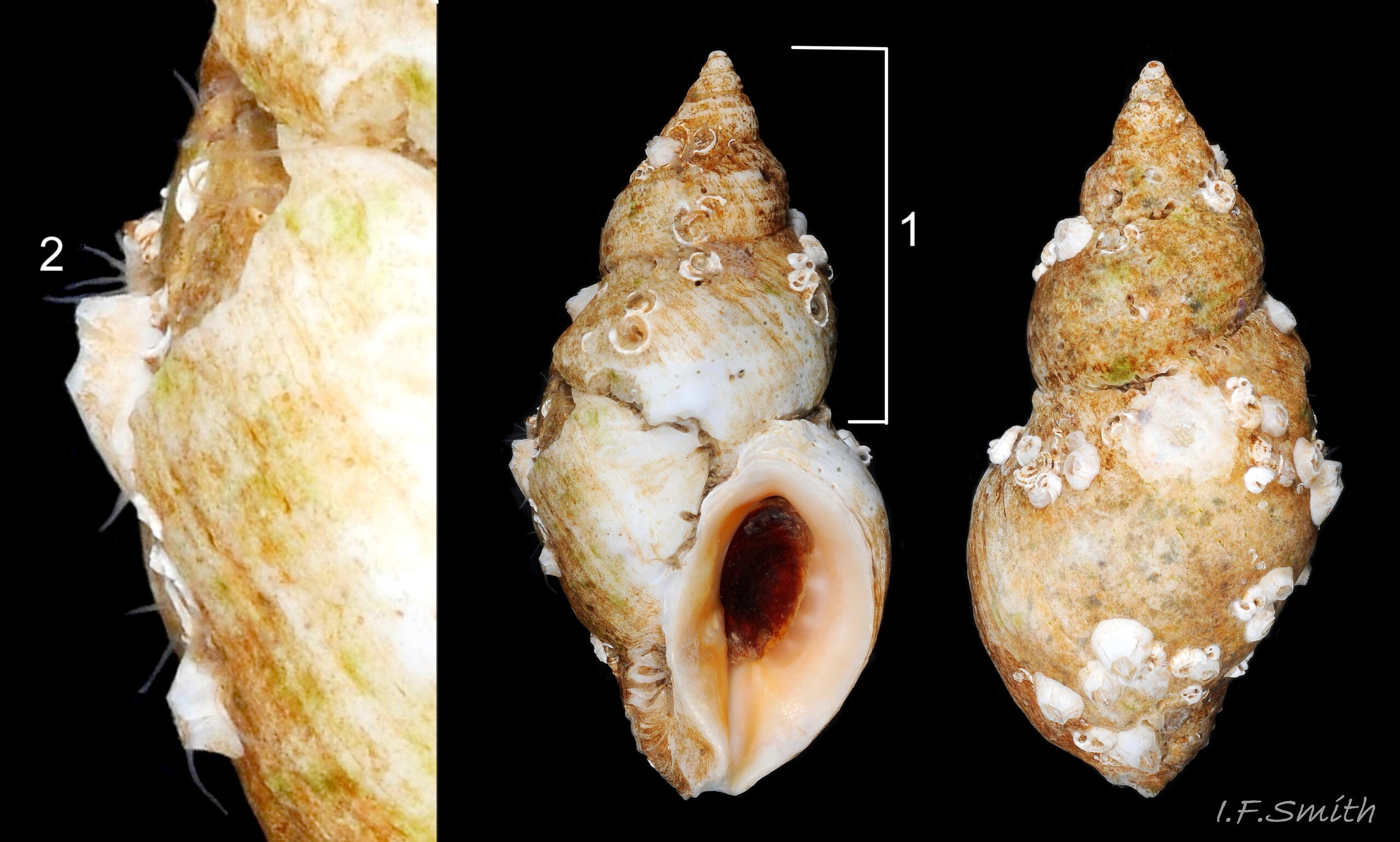
01 Nucella lapillus

02 Nucella lapillus
Squat, extremely exposed shore form with short spire height (1) 24% of shell height. Large aperture 76% of shell height. Fourteen well developed flattened, spiral ridges on body-whorl. Shell height 25 mm, breadth 17.4 mm. West coast of Hoy, Orkney (Atlantic …

03 Nucella lapillus. Menai Strait, Wales. February 2013.

04 Nucella lapillus. Lleyn Peninsula, Wales. September 2009.

05 Nucella lapillus

06 Nucella lapillus. Height 15mm. Menai Strait, Wales. February 2013.

07 Nucella lapillus. Menai Strait, Wales. September 2011.

08 Nucella lapillus

09 Nucella lapillus. Great Orme, Wales. July 2011.

10 Nucella lapillus. Height 30.6 mm. Mouth of Mersey Estuary, England. July 2014

11 Nucella lapillus. Kincardineshire, N.E. Scotland. September 1970.

12 Nucella lapillus. Height 30mm. Mouth of Mersey Estuary, England. July 2014.

13 Nucella lapillus. Juveniles 2.2 mm to 3 mm high. Menai Strait, Wales. August 2013.

14 Nucella lapillus. Height 12 mm. Menai Strait, Wales. February 2013.

15 Nucella lapillus. Height 25.8mm. Mouth of Dee Estuary, Cheshire, England.

16 Nucella lapillus. Height 34.1mm. Meirionnydd, Wales. July 1969.

17 Nucella lapillus. Height 18.7mm. Cornwall, England. August 1982. Coll. J. Light.

18 Nucella lapillus. Height 32mm. N. Devon, England. April 1985. Coll. J. Light.

19 Nucella lapillus. Lleyn Peninsula, Wales. September 2009.

20 Nucella lapillus. Operculum 9.5mm long from adult with shell 40.6mm high. Menai Strait, Wales. June 2013.

21 Nucella lapillus. Menai Strait and (right) Great Orme, Wales.
22 Nucella lapillus. Shell height 32.5mm. Mouth of Mersey Estuary, England. July 2014.
23 Nucella lapillus. Height 15mm. Menai Strait, Wales. February 2013.
24 Nucella lapillus. Menai Strait, Wales. September 2011.
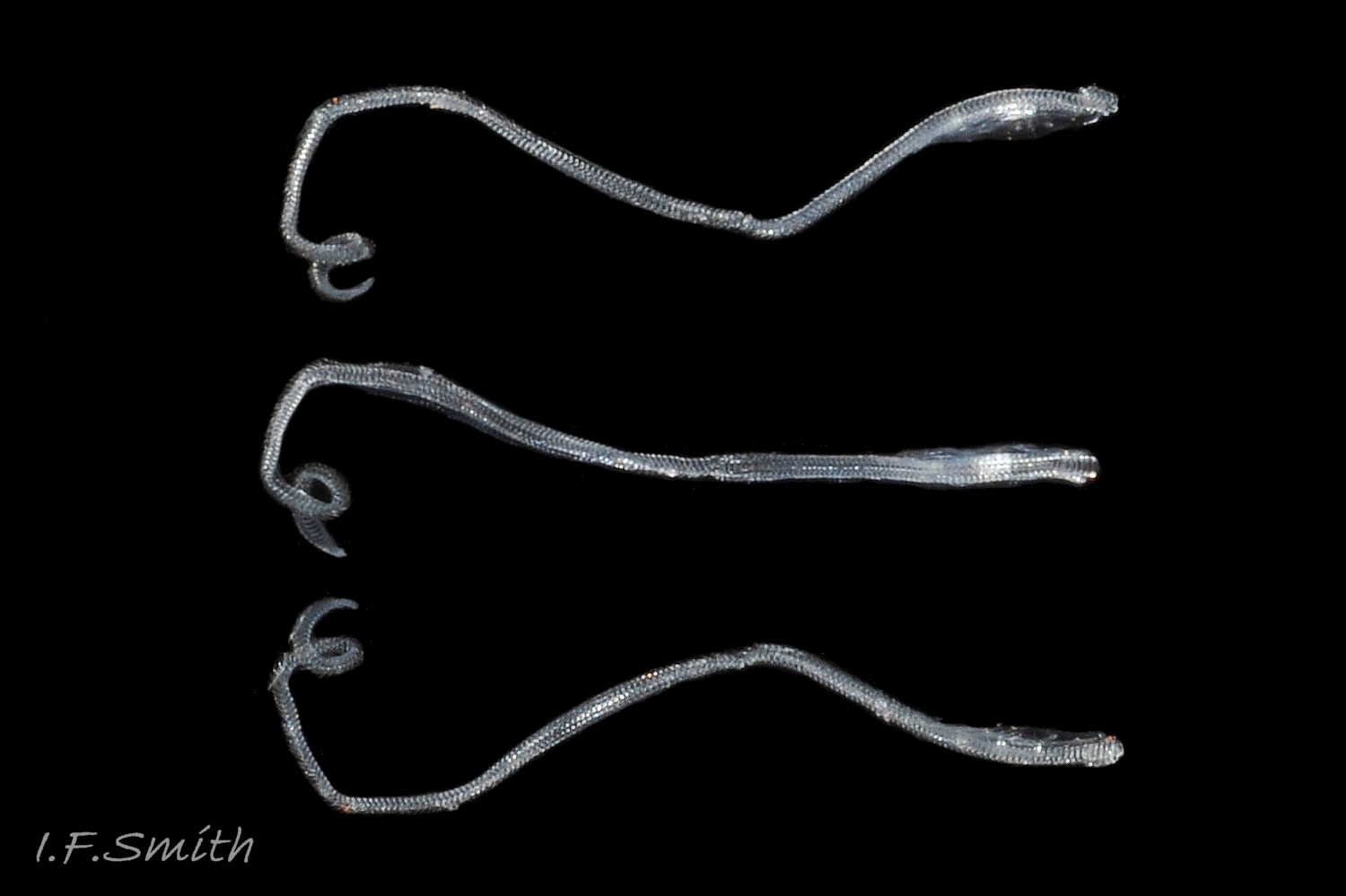
25 Nucella lapillus. Mouth of Mersey Estuary, England. July 2014.

26 Nucella lapillus. Shell height 27mm. Mouth of Mersey Estuary, England. July 2014.
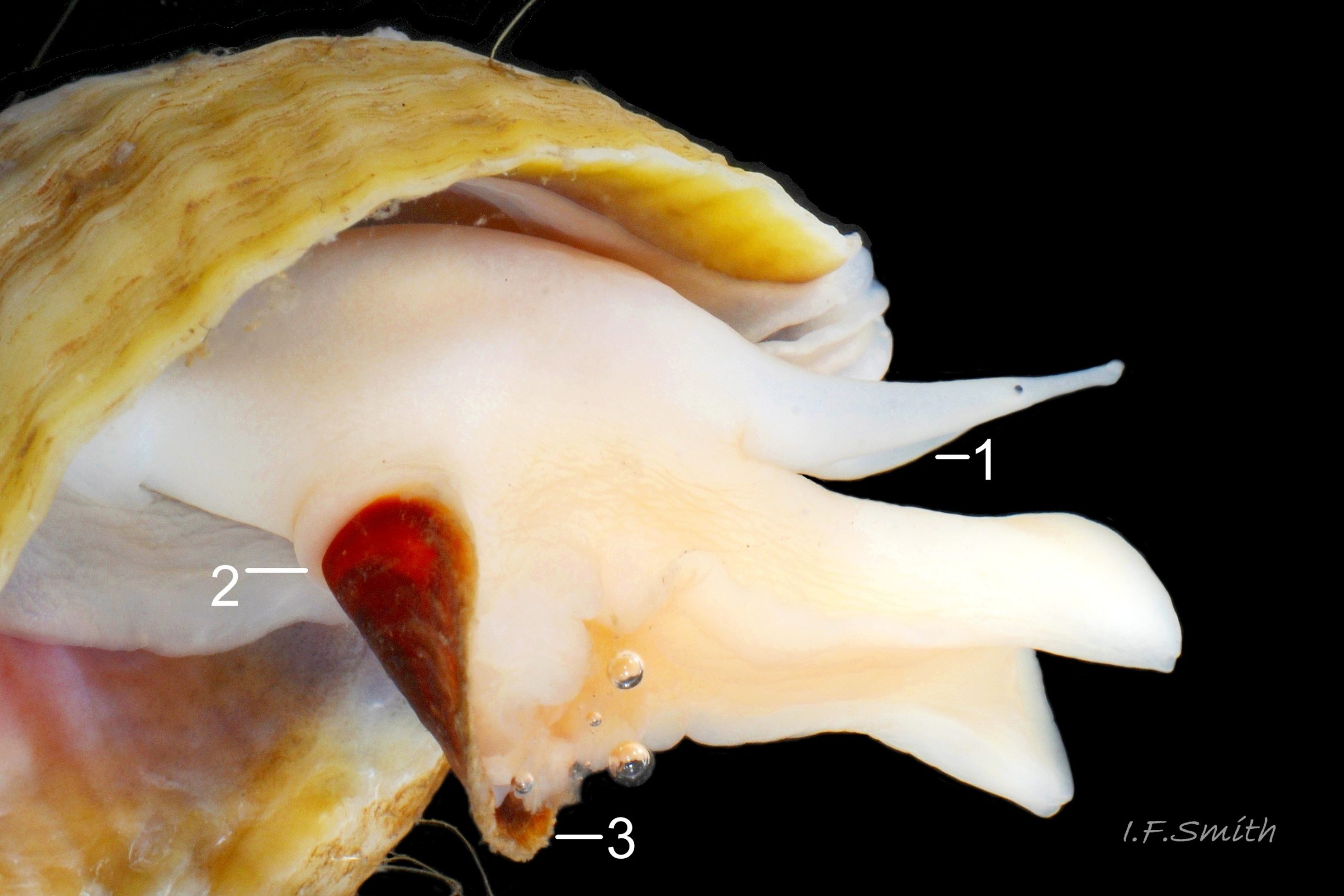
27 Nucella lapillus. Shell height 32.5mm. Mouth of Mersey Estuary, England. July 2014.

28 Nucella lapillus. Menai Strait, Wales. September 2011.

29 Nucella lapillus. Great Orme, Wales. July 2011.

30 Nucella lapillus. Menai Strait, Wales. September 2011.

31 Nucella lapillus. Juvenile. Shell height 3mm. Menai Strait, Wales. August 2013.

32 Nucella lapillus. Great Orme, Wales. July 2011.

33 Nucella lapillus. Juvenile. Shell height 3mm. Menai Strait, Wales. August 2013.
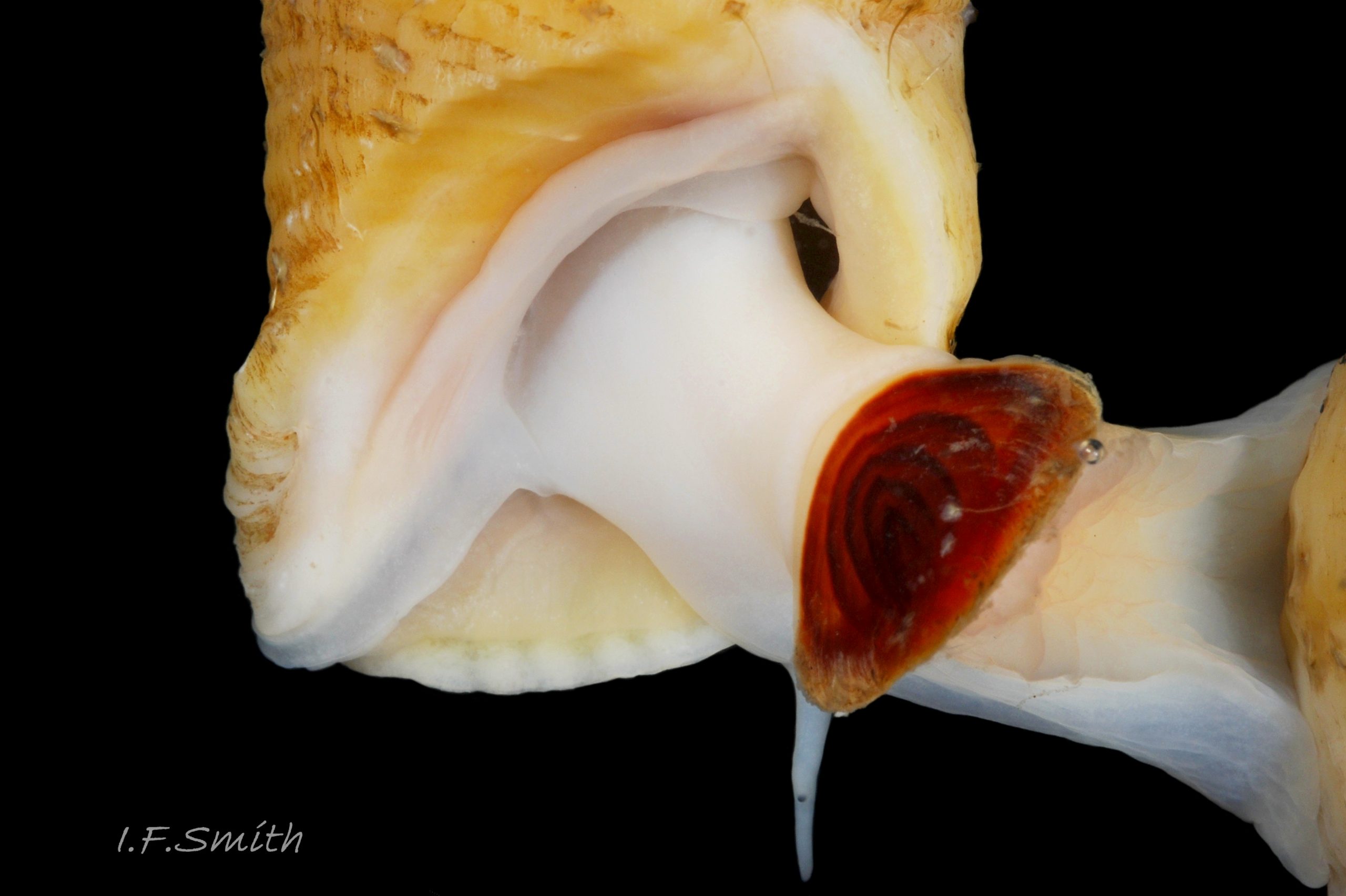
34 Nucella lapillus. Shell height 31.4mm. Mouth of Mersey Estuary, England. July 2014.
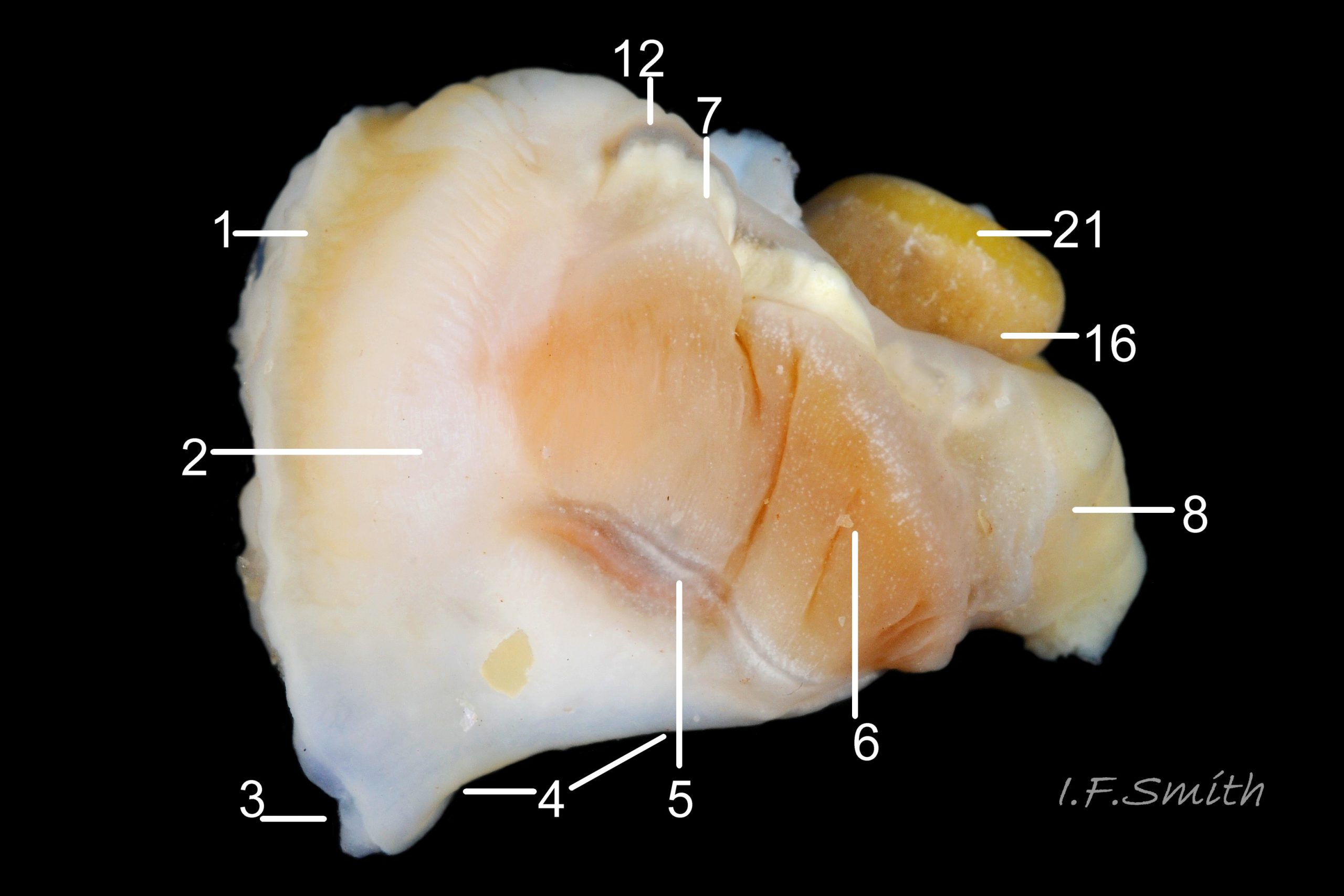
35a Nucella lapillus. Male removed from shell, dorsal view. Mouth of Mersey Estuary, England. July 2014.
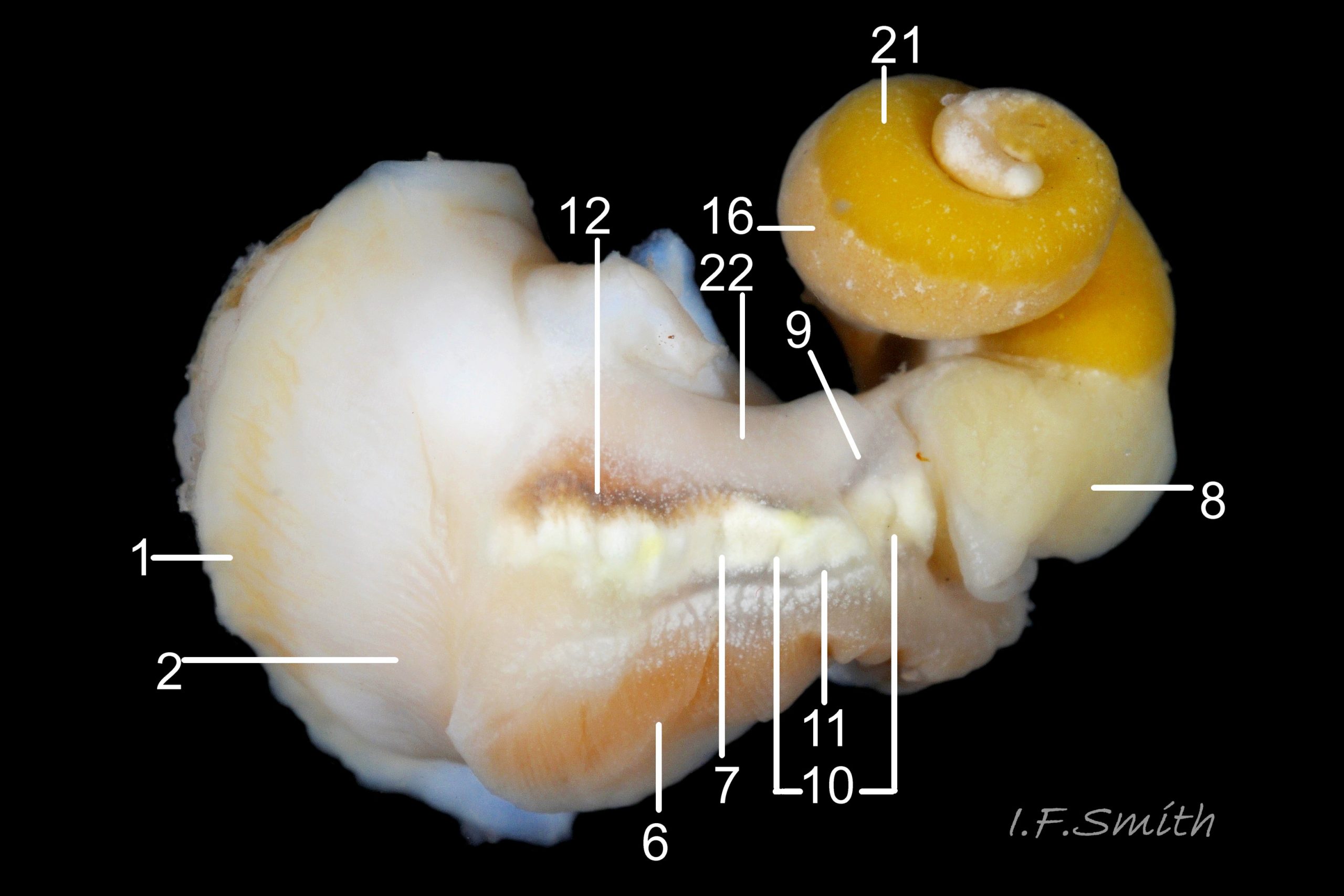
35b Nucella lapillus. Male removed from shell, tilted to expose right side. Mouth of Mersey Estuary, England. July 2014.
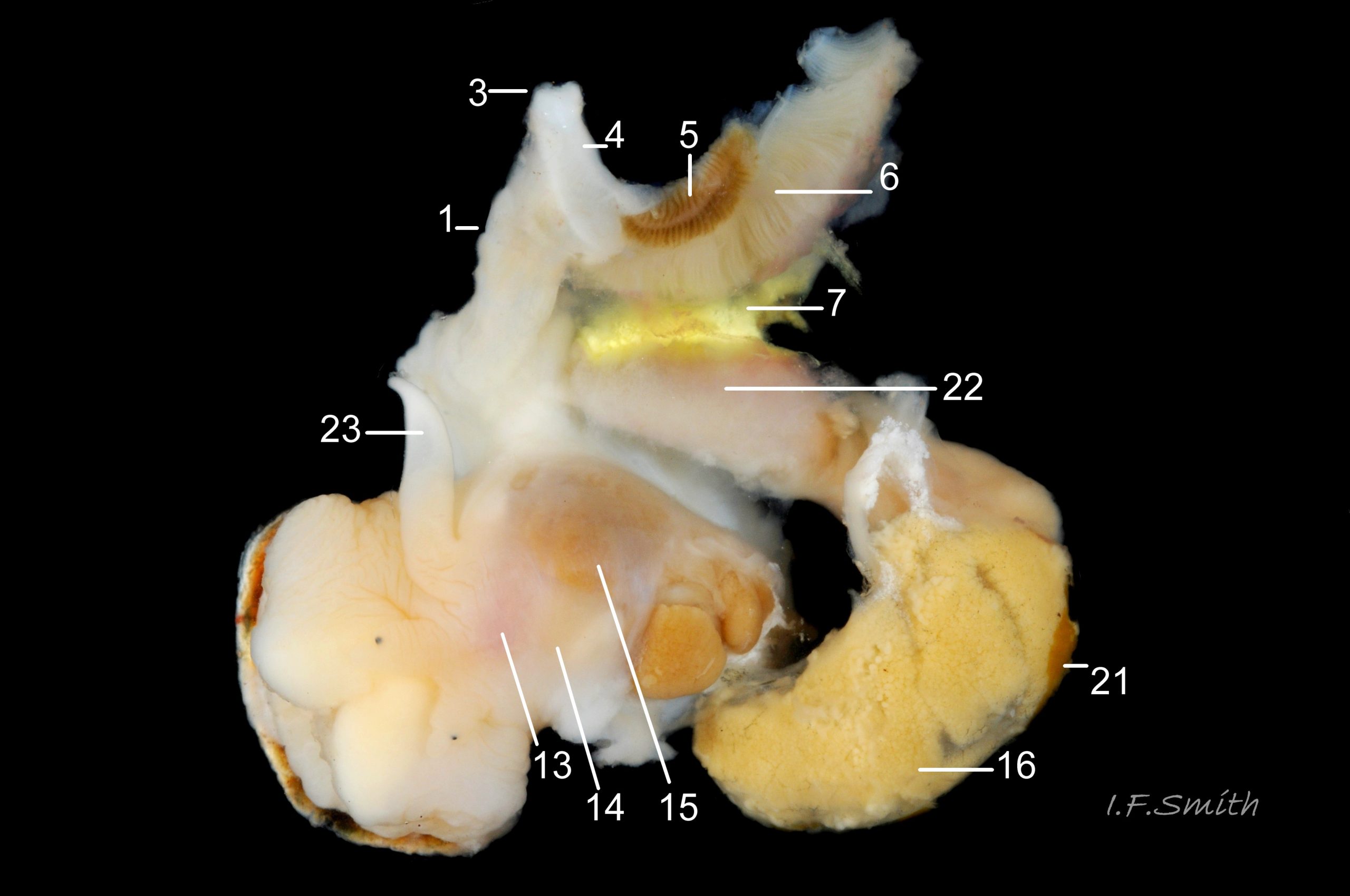
36 Nucella lapillus. Male removed from shell; mantle cut on left and turned out to animal’s right. Mouth of Mersey Estuary, England. July 2014.
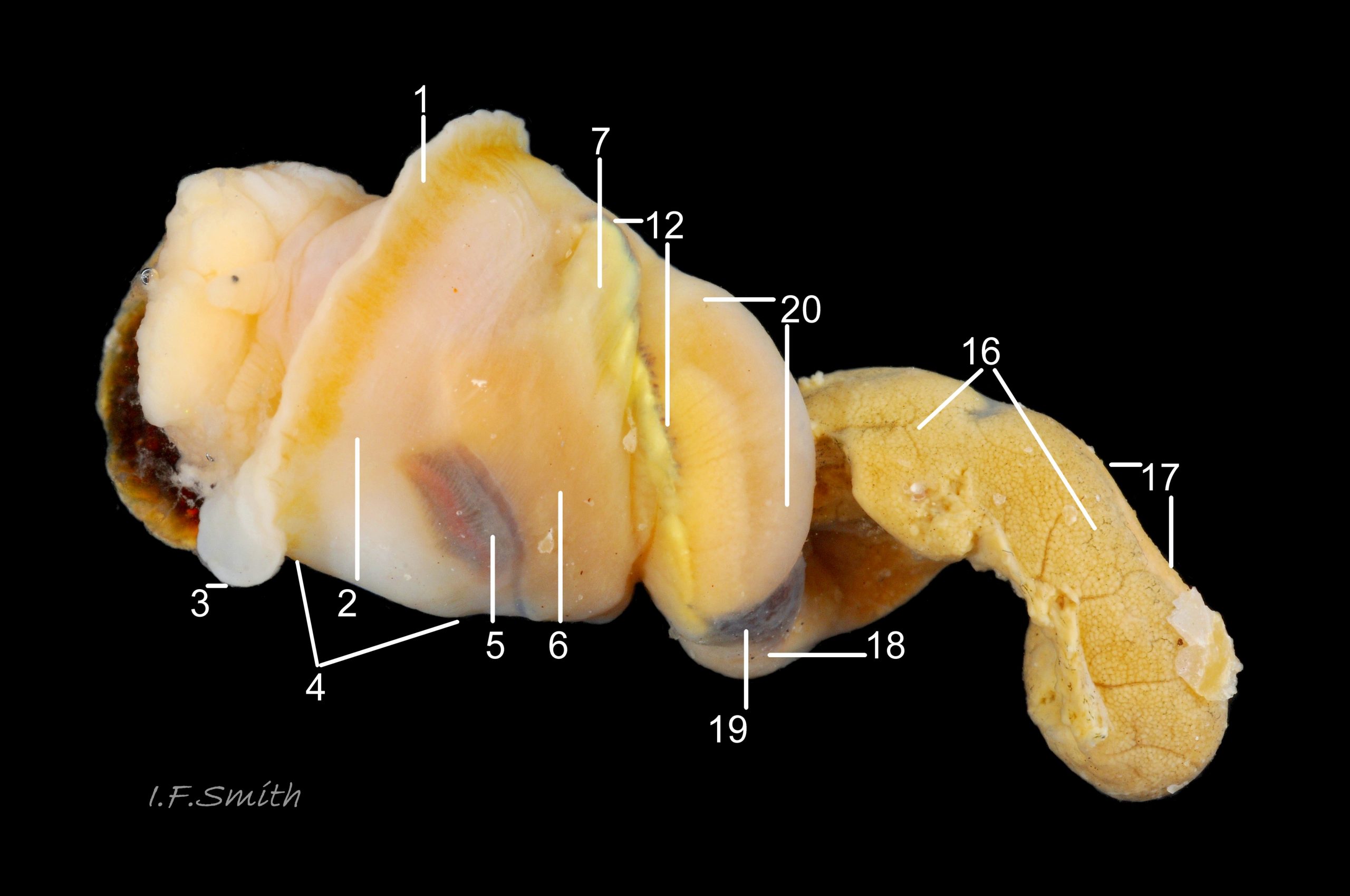
37 Nucella lapillus. Female removed from shell. Mouth of Mersey Estuary, England. July 2014.
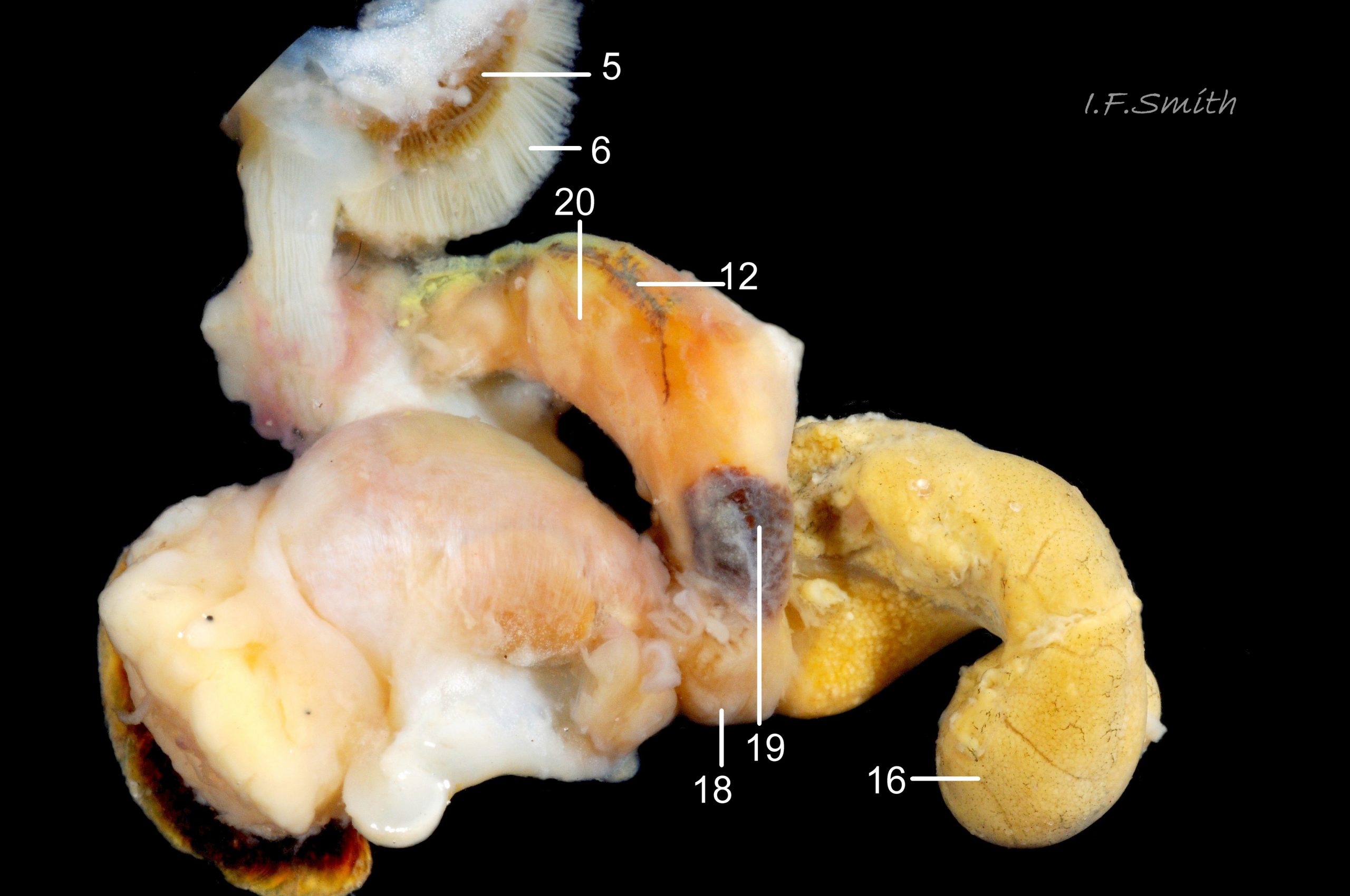
38 Nucella lapillus. Female removed from shell, and mantle cut on left and turned out to animal’s right. Mouth of Mersey Estuary, England. July 2014.

39 Nucella lapillus 31mmhigh (centre), Urosalpinx cinerea 30mm high (left), & Buccinum undatum 52mm high (right).

39a Nucella lapillus. Feeding on Mytilus edulis. North Cornwall, England. April 2008. Leg. & copyright N. Ward.
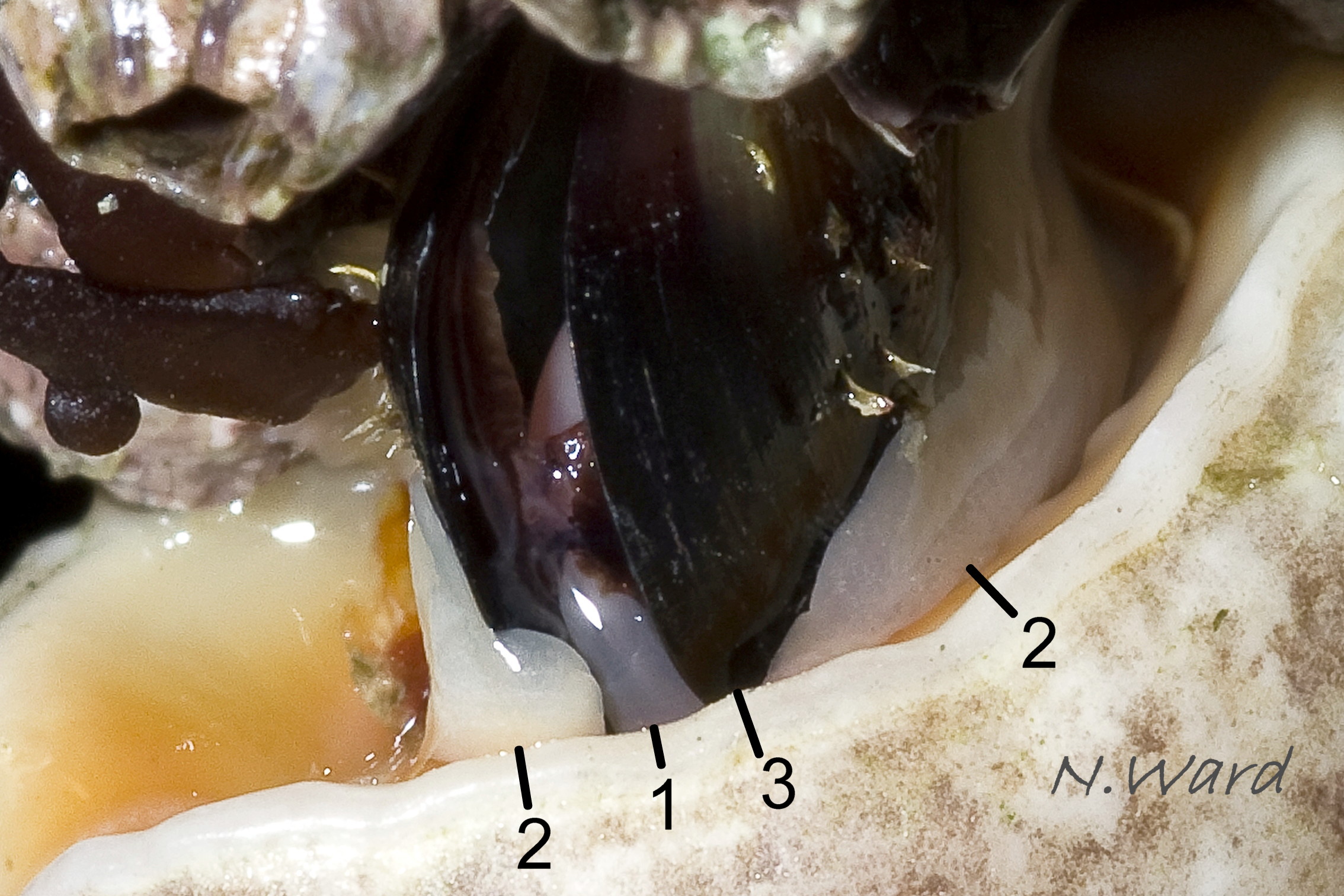
39b Nucella lapillus. Feeding on Mytilus edulis. North Cornwall, England. April 2008. Leg. & copyright N. Ward.

40 Nucella lapillus. Menai Strait, September 2009.

40a Nucella lapillus boring into Patella depressa (33mm long). Lleyn, North Wales. September 2015.
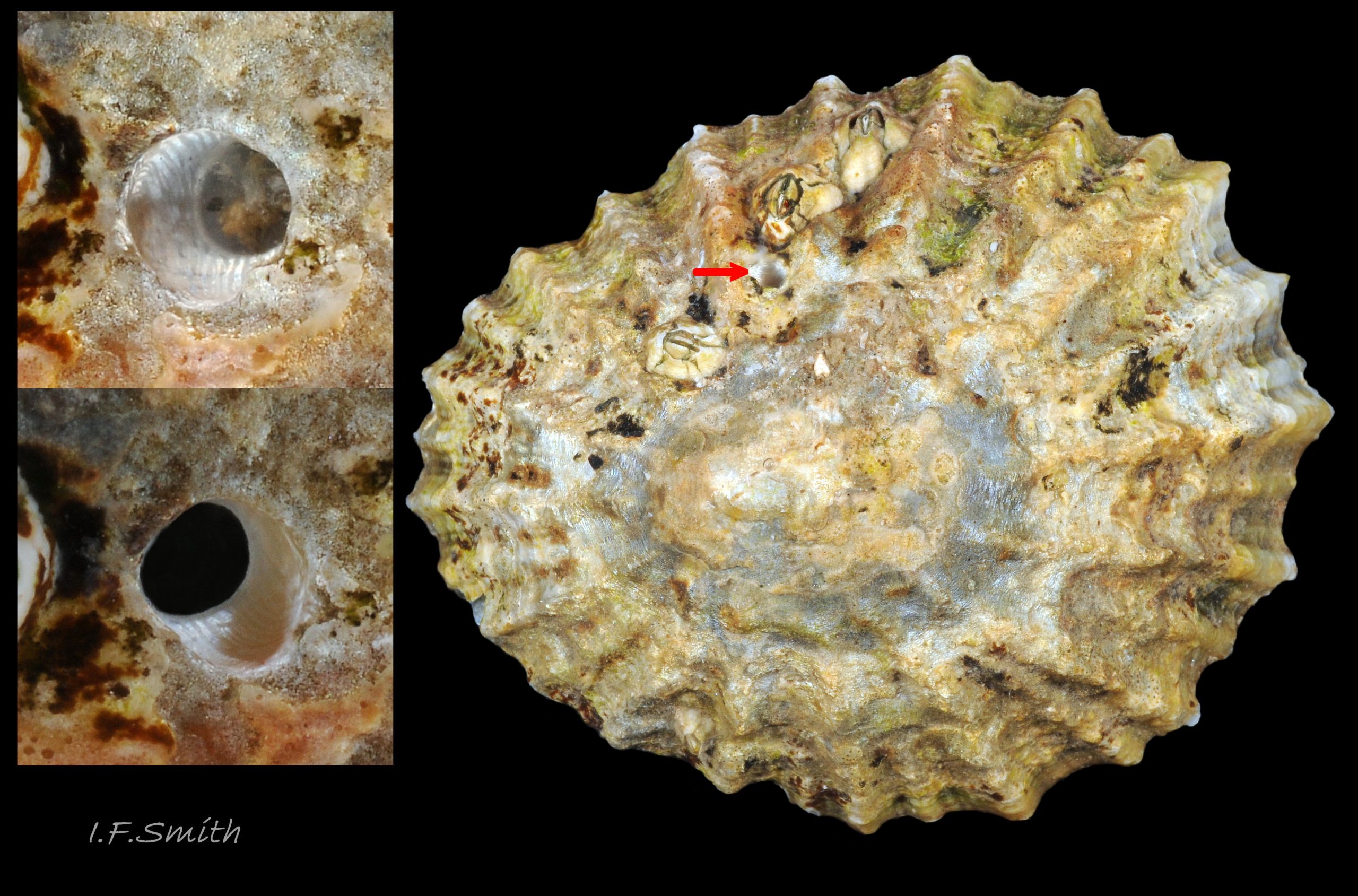
40b Nucella lapillus bore-hole on Patella depressa (33mm long). Lleyn, North Wales. September 2015.

40c Nucella lapillus incomplete bore-hole on Patella depressa (29.5mm long). Lleyn, North Wales. September 2015.
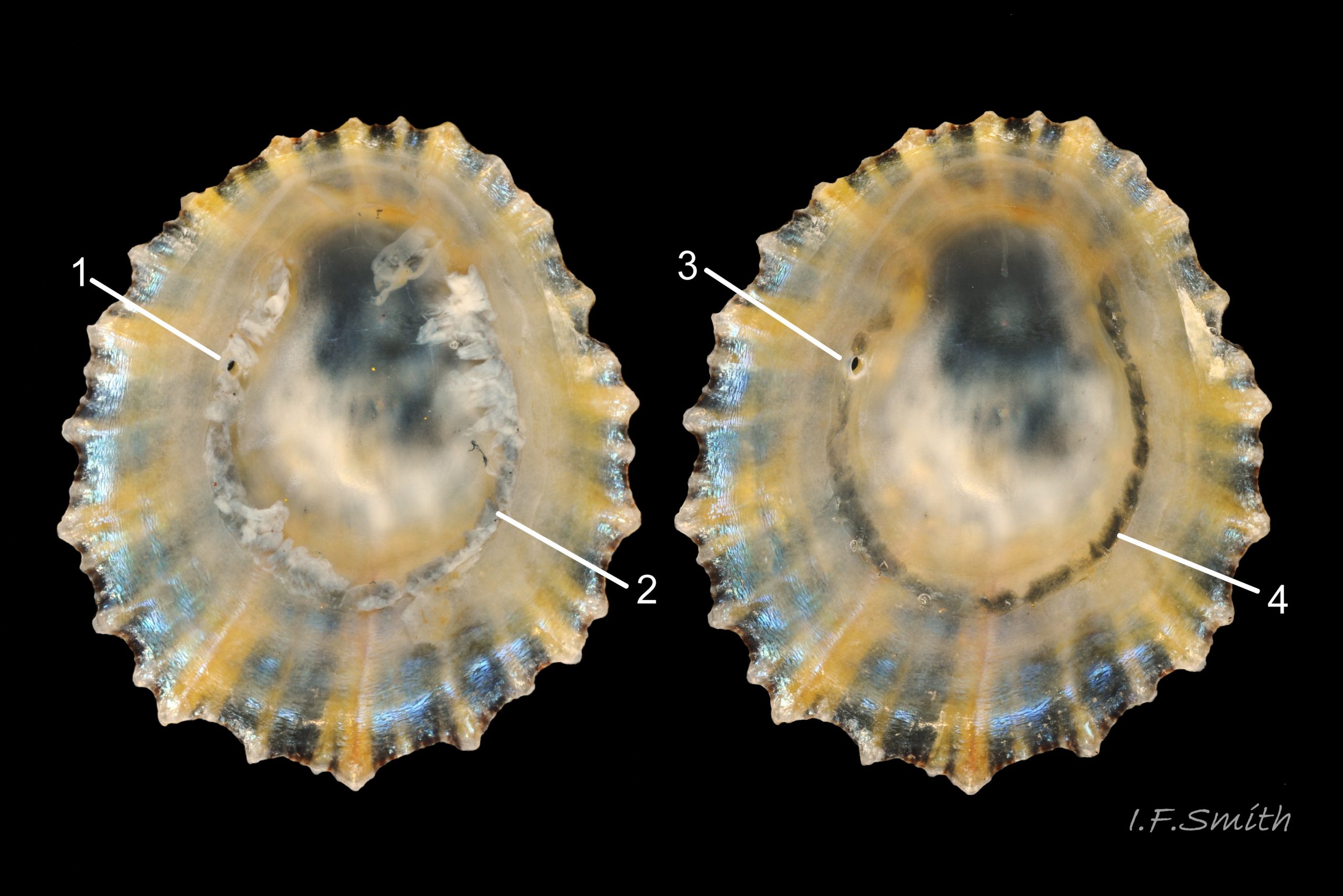
40d Nucella lapillus bore-hole viewed from interior of Patella depressa (33mm long). Lleyn, North Wales. September 2015.

40e Nucella lapillus album; Patella shells bored by Nucella lapillus. Lengths 39.7mm, 31.9mm and 36.6mm. Guernsey. Leg. D. Flint.

41 Nucella lapillus. Entrapped by Mytilus. Mouth of Mersey Estuary, England. July 2014.

41a Nucella lapillus album. Prey species, Mytilus edulis. Juvenile 11.3mm long. Mouth of Mersey Estuary, England. July 2014.

42 Nucella lapillus. 25mm high & 30.7mm high. From extremely exposed shore and extremely sheltered shore (6km apart) Hoy, Orkney, Scotland. July 1973.

43 Nucella lapillus. Egg capsules. Menai Strait, Wales. March 2010.
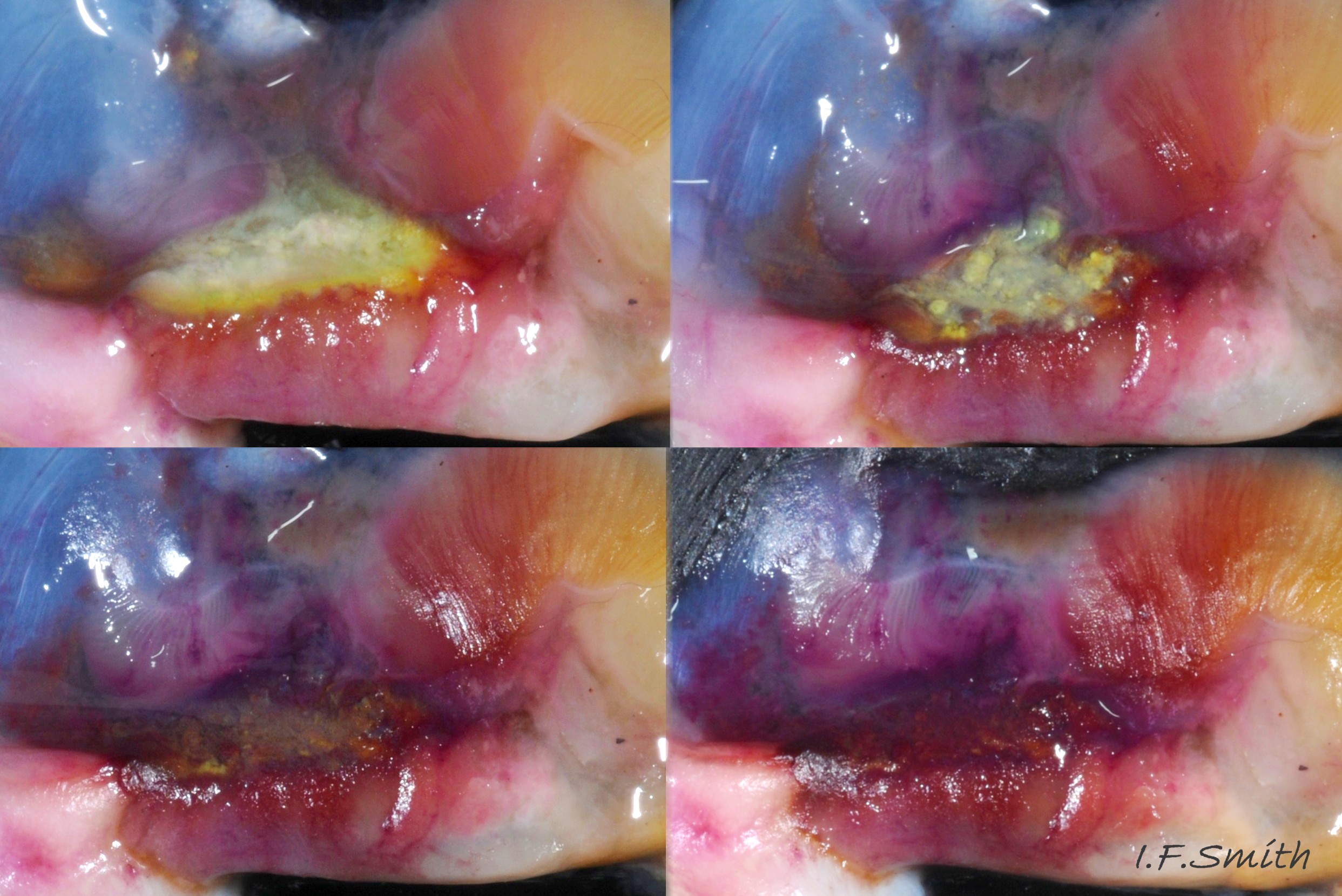
44 Nucella lapillus. Hypobranchial gland exposed to light and air.
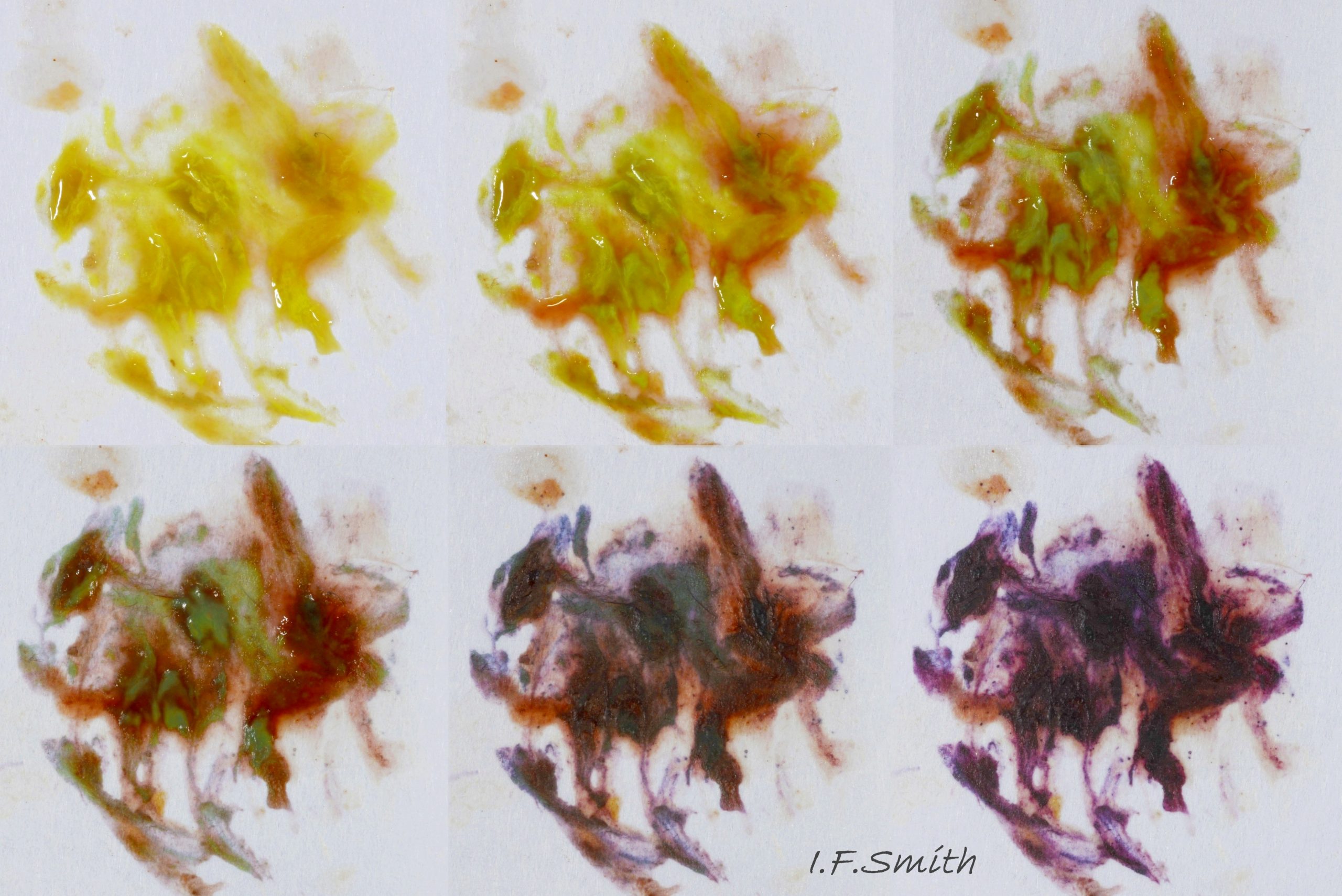
45 Nucella lapillus. Hypobrachial gland mucus on paper.
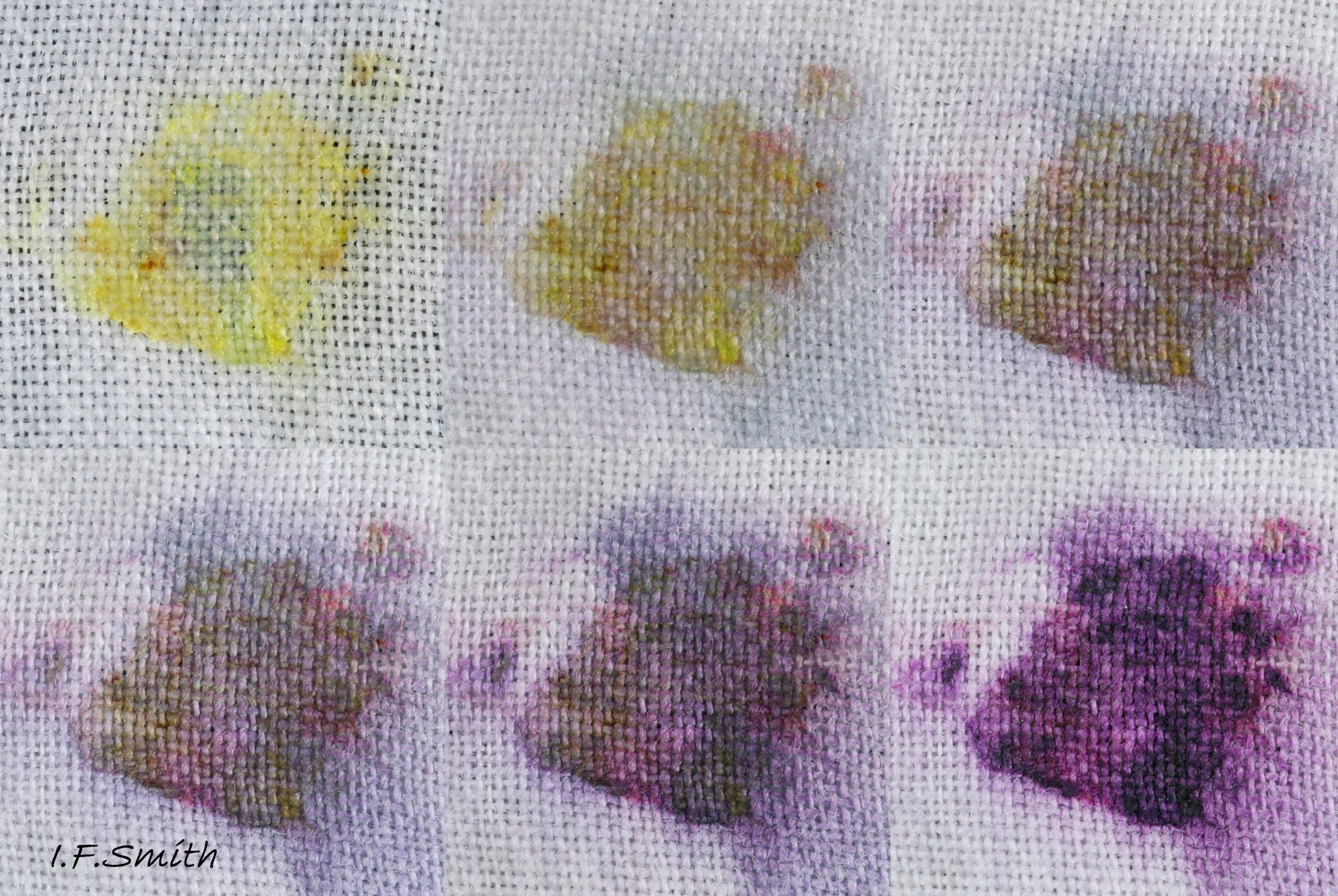
46 Nucella lapillus. Hypobrachial gland mucus on cloth.
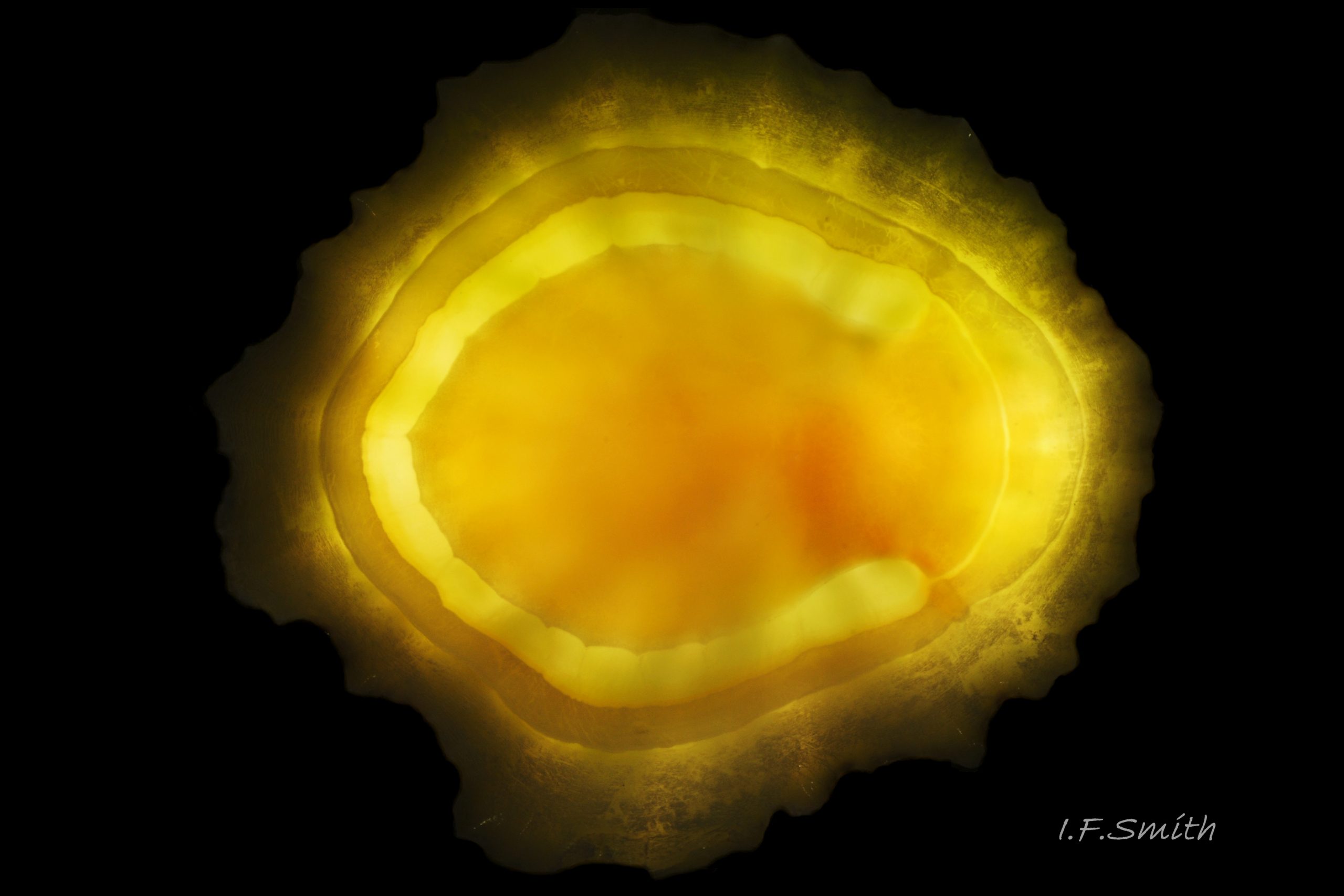
47 Nucella lapillus album. Backlit image of interior of Patella ulyssiponensis shell. Lleyn, North Wales. September 2015.
Nucella lapillus (Linnaeus, 1758)
Current taxonomy: World Register of Marine Species (WoRMS)
www.marinespecies.org/aphia.php?p=taxdetails&id=140403
Synonyms: Purpura lapillus (Linnaeus, 1758); Thais lapillus (Linnaeus, 1758); Polytropa lapillus (Linnaeus, 1758);
Vernacular: Dog whelk; Dog winkle; Purple; Rock whelk; Horse winkle (English); Gwichiad/Chwalcen y cŵn; Cragen foch felen (Welsh); Purpursnegl (Danish & Norwegian); purperslak (Dutch); Pourpre petit pierre; pourpre (French); Nordische Steinchenschneke/Purpurschneke (German); Purpur snäcke (Swedish);
Illustrated pdf of account available at
www.researchgate.net/profile/Ian_Smith19/contributions
GLOSSARY BELOW
Shell Description
Adults usually 17 to 30mm high, up to 45mm littorally 01 Nucella lapillus. and over 60mm sublittorally. Shape varies from long and slender (spire 55% of shell height) 05 Nucella lapillus to short and squat (spire10%, body-whorl 90% of shell-height) 02 Nucella lapillus.. Apex slightly twisted, sharp if uneroded. Whorls moderately convex. Sutures usually slight, may be deeper near apex 03 Nucella lapillus , can be almost obliterated by erosion 04 Nucella lapillus. Aperture height varies greatly, about 45% 05 Nucella lapillus to 80% 02 Nucella lapillus of shell height. Juveniles and sub-adults (less than c17mm high) have thin shell- walls with sharp outer lip 06 Nucella lapillus. Adults (when 17mm or more high) usually cease longitudinal growth and develop thick shell-walls and line of blunt bosses within outer lip of aperture 07 Nucella lapillus Sometimes more than one row of bosses if growth arrested prematurely. Outer lip smooth, interior slightly shiny, not sharp, but tapers from bosses to edge. Aperture rapidly constricts into throat 08 Nucella lapillus. Adapical angle acute when young 06 Nucella lapillus , rounded as shell thickens 09 Nucella lapillu. Short, straight, open siphonal canal at base of aperture. Columellar and parietal lips are wide, smooth and slightly shiny 10 Nucella lapillus ; columellar lip overhangs siphonal canal slightly; parietal lip continuous with outer lip through the adapical angle. Siphonal fasciole pronounced on some 11 Nucella lapillus. Sculpture of flattened, spiral ridges (about 12 on body-whorl) 12 Nucella lapillus & 02 Nucella lapillus , often indistinct or absent 09 Nucella lapillus. Growth lines usually indistinct. Newly hatched young have globular, smooth, glossy shell (protoconch) of 2½ whorls about 1mm high. Early growth to c.3mm very often has prominent imbricate sculpture 13 Nucella lapillus which, with the protoconch, may persist on early whorls of uneroded adults 03 Nucella lapillus and occasionally continue on later whorls 14 Nucella lapillus & 15 Nucella lapillus. Ground colour white 07 Nucella lapillus, whitish-grey 04 Nucella lapillus, yellow 12 Nucella lapillus , orange 16 Nucella lapillus, brown to black 17 Nucella lapillus, or mauve to pink 03 Nucella lapillus. Interior of aperture shows exterior colours on thin translucent young shells 06 Nucella lapillus, and thickened adult shells are often coloured internally 09 Nucella lapillus, 10 Nucella lapillus & 12 Nucella lapillus. Sometimes exterior colours combined in spiral bands; narrow and confined to grooves between spiral ridges 18 Nucella lapillus. , or broad 06 Nucella lapillus Growths 07 Nucella lapillusand erosion 04 Nucella lapillus can affect colour. Light-horn coloured periostracum often on small juveniles 13 Nucella lapillus, absent or insignificant on adults. Broad, angular, mahogany, non-spiral operculum 20 Nucella lapillus; thin outer face extruded from transverse groove at anterior of opercular disc has many fine, gently-curving growth lines increasing in length away from nucleus on posterior (labial) edge; thick inner face made of stack of annual growth plates (adventitious layers extruded from circles of glands on opercular disc) with excentric nucleus and matt surface where opercular disc attached (termination of columellar muscle). Posterior (labial) margin beyond disc attachment (c.25% of total operculum surface) covered by shiny varnish from transverse groove across posterior of opercular disc. Operculum nearly opaque on old adults, but younger ones have translucent operculum through which adventitious plates show as strong concentric rings that obscure fine exterior growth lines 21 Nucella lapillus.
Body Description
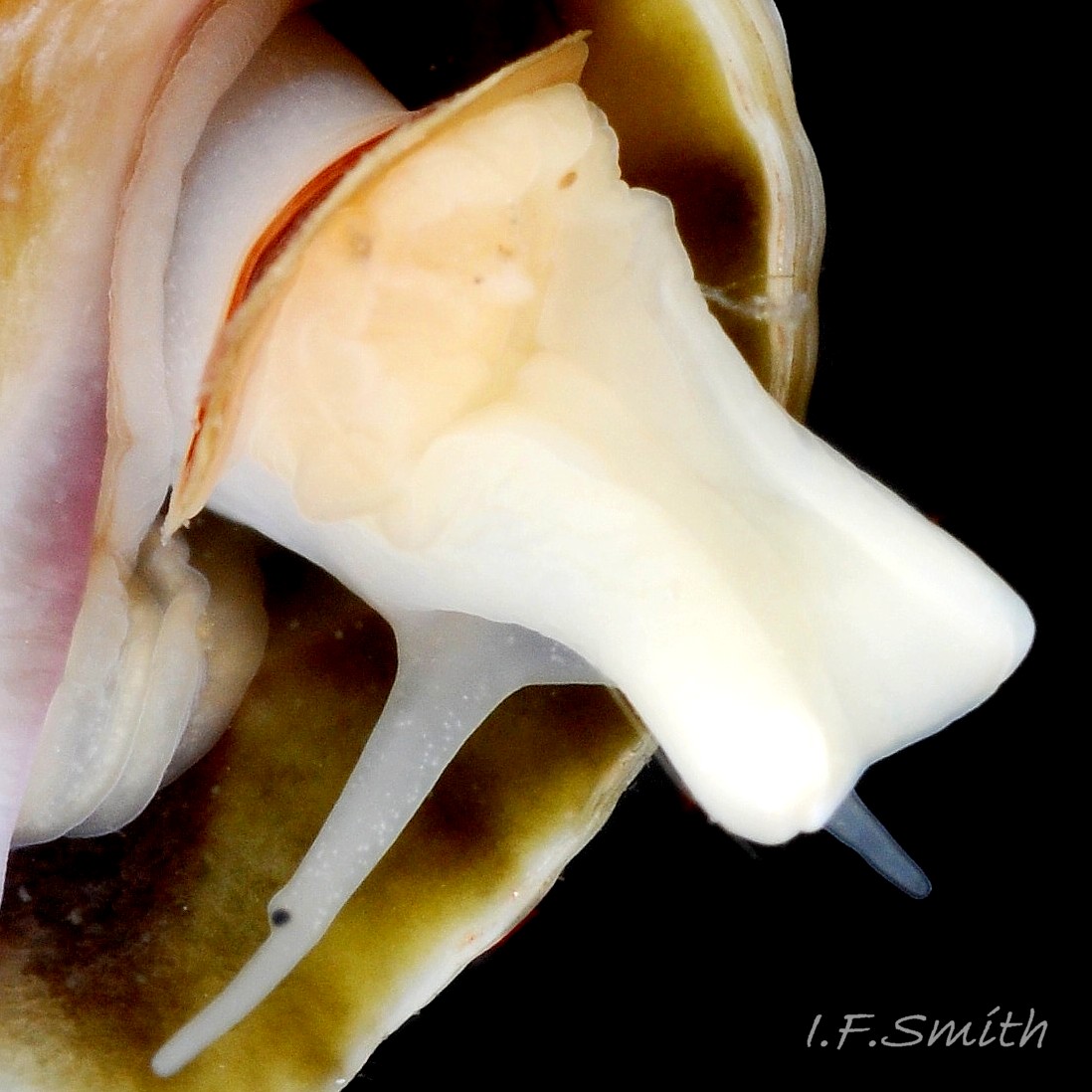
Flesh, apart from mantle, entirely pure-white or yellowish-white, variably translucent 22 Nucella lapillus , sometimes opaque white freckles visible on surface of more translucent parts 23 Nucella lapillus. Head consists of transverse ridge at juncture of cephalic tentacles. No snout visible, except, when feeding, a short, thin proboscis 39b Nucella lapillus is everted like a sock turned inside-out from opening at base of head’s anterior 24 Nucella lapillus . Pink buccal mass 22 Nucella lapillus with narrow, short radula, about 30% of shell length; only three teeth per row 25 Nucella lapillus. Long, tapering, rounded cephalic tentacles; translucent white, sometimes with opaque white flecks 23 Nucella lapillus. Basal two thirds of cephalic tentacle thickened by fusion with eye peduncle; black eye on summit of outer face of thickened section 26 Nucella lapillus; divided ventrally on one examined specimen 27 Nucella lapillus . Mantle edge thickened, usually yellowish even on pure white animals 28 Nucella lapillus, occasionally with a little brown. Mantle folded into white inhalent siphon on left of body, continues as channel to osphradium and substantial buff ctenidium within mantle cavity; siphon extends only slightly beyond end of siphonal canal of shell 29 Nucella lapillus Faint pink of myoglobin-rich buccal mass, white salivary glands and other internal organs sometimes visible behind head of well-extended animal 22 Nucella lapillus. Flat, strap-like (when live) penis attached sub-dorsally behind right tentacle; recurved up to 360° 24 Nucella lapillus, may be pulled straighter during emergence of body from shell 30 Nucella lapillus. Foot can change between oblong 31 Nucella lapillus and bluntly oval 32 Nucella lapillus, often constricted behind propodium. Anterior edge bilaminate 33 Nucella lapillus, sole and dorsum coloured as body. Small opercular disc does not extend beyond or overlap edge of operculum 24 Nucella lapillus, apart from anterior edge of operculum being exuded from opercular groove 27 Nucella lapillus. Strong white columellar muscle attaches opercular disc to columella of shell 34 Nucella lapillus. Sole of foot divided along midline; most obvious when foot folds along it 23 Nucella lapillus. Sucker-like accessory boring organ (ABO) in sac within foot on midline near anterior; when everted swells into a large proboscis-like projection. Female has ventral pedal gland for moulding egg capsules, anterior to centre of sole.
Internal anatomy
When extracted from shell, organs in place but indistinctly seen through translucent mantle on anterior half of body. When mantle cut along pallial inhalent channel and opened out to the right of the animal like a page of a book, organs attached to its inner face are displaced but more clearly visible.
KEY to items on images 35a Nucella lapillus & 35b Nucella lapillus (extracted male), 36 Nucella lapillus (opened male) 37 Nucella lapillus (extracted female) 38 Nucella lapillus. (opened female). If key is read in order, the functions and inter-relationship of the organs may be understood.
Mantle
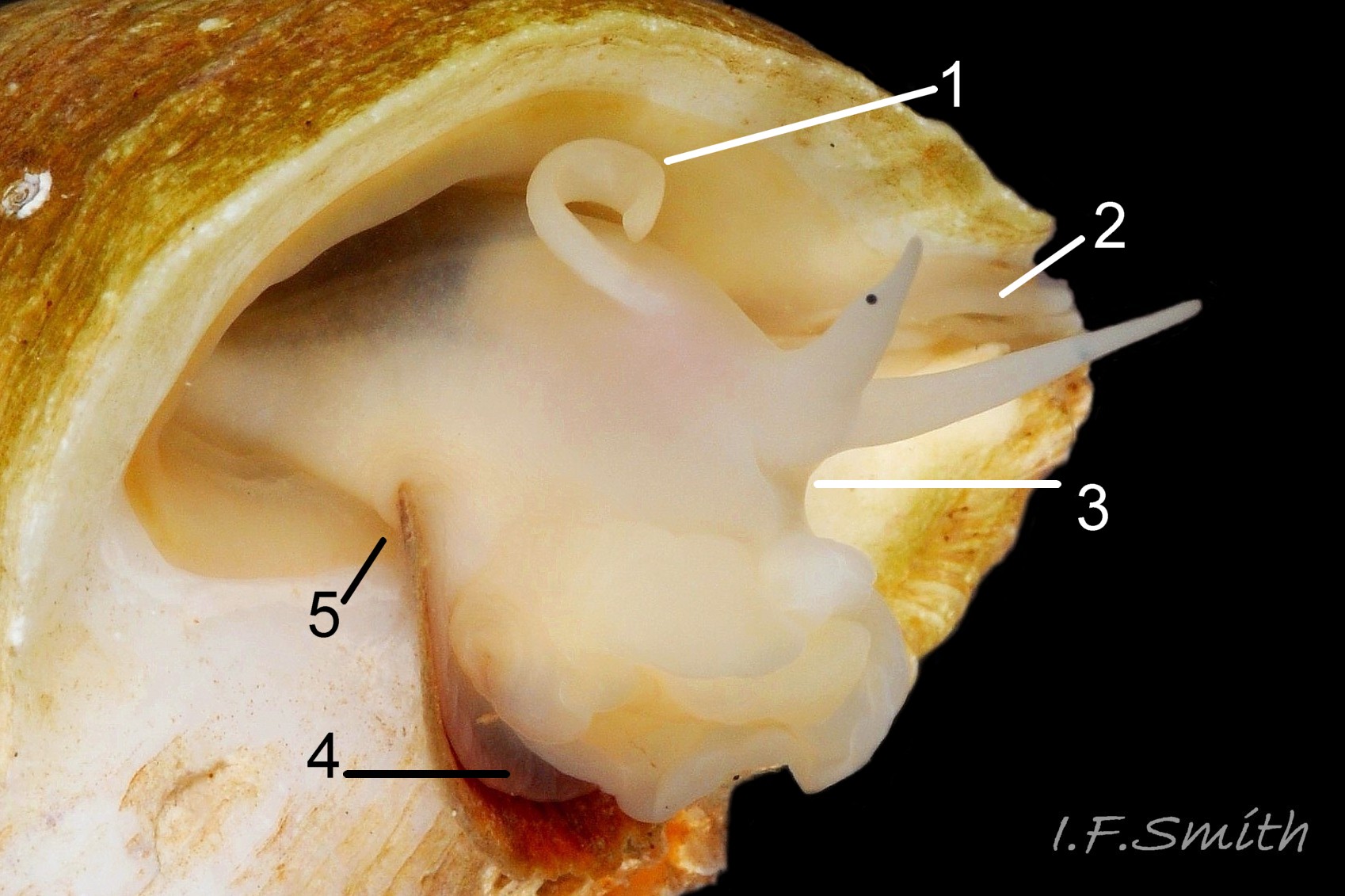
1: mantle edge – substantial, opaque, yellowish, slightly flounced, anterior border of mantle skirt. Only part that produces exterior layers of shell. Sometimes flounces exaggerated and make protruding shell-growth that forms imbricate sculpture. (Images 35a&b, 36, 37)
2: mantle skirt – forms roof of mantle cavity and, when uncut, partially obscures organs within. (Images 35a&b, 37)
Respiratory features
3: siphon – white, folded extension of mantle that rests in shell’s siphonal canal and draws in inhalent current of water when animal extended. (Images 35a, 36, 37)
4: pallial inhalent channel – distinct, white fold in mantle forming a channel from siphon to osphradium. (Images 35a, 36, 37)
5: osphradium – dark, bipectinate chemo-receptor at inner end of inhalent channel that tests water approaching ctenidium for quality and scents of food, prey, predators and/or mates. So highly developed on Nucella that Jeffreys (1867) thought it was second ctenidium. (Images 35a, 36, 37, 38)
6: ctenidium – substantial, buff-white gill with many fine leaflets that receive oxygen from inhalent water, and oxygenate blood passing through them. (Images 35a&b, 36, 37, 38)
7: hypobranchial gland – puckered gland that produces mucous to trap particles from inhalent water before reaches ctenidium, and to transport particles out of mantle cavity. Other functions might be attraction of mates by scent, and acrid smell/taste to discourage predators. Initially cream-white, changing to yellow, green, purple etc when exposed to light and air. (Images 35a&b, 36,37)
Vascular features
8: kidney – de-oxygenated blood has urea etc. removed and excreted by kidney before passing to efferent renal vein [9]. (Images 35a&b)
9: efferent renal vein – carries blood away from kidney. (Image 35b)
10: Hypobrachial vessels – carry blood from efferent renal vein through hypobranchial gland to afferent vessel of ctenidium [11]. (Image 35b)
11: afferent vessel of ctenidium – receives blood from hypobranchial vessels and passes it into leaflets of adjacent ctenidium for oxygenation. (Image 35b)
12: rectal gland – long, linear gland; dark brown to purple-black in adults. Function uncertain, perhaps produces substances that supplement the excretory activity of the kidney. (Images 35a&b, 37, 38)
Alimentary features
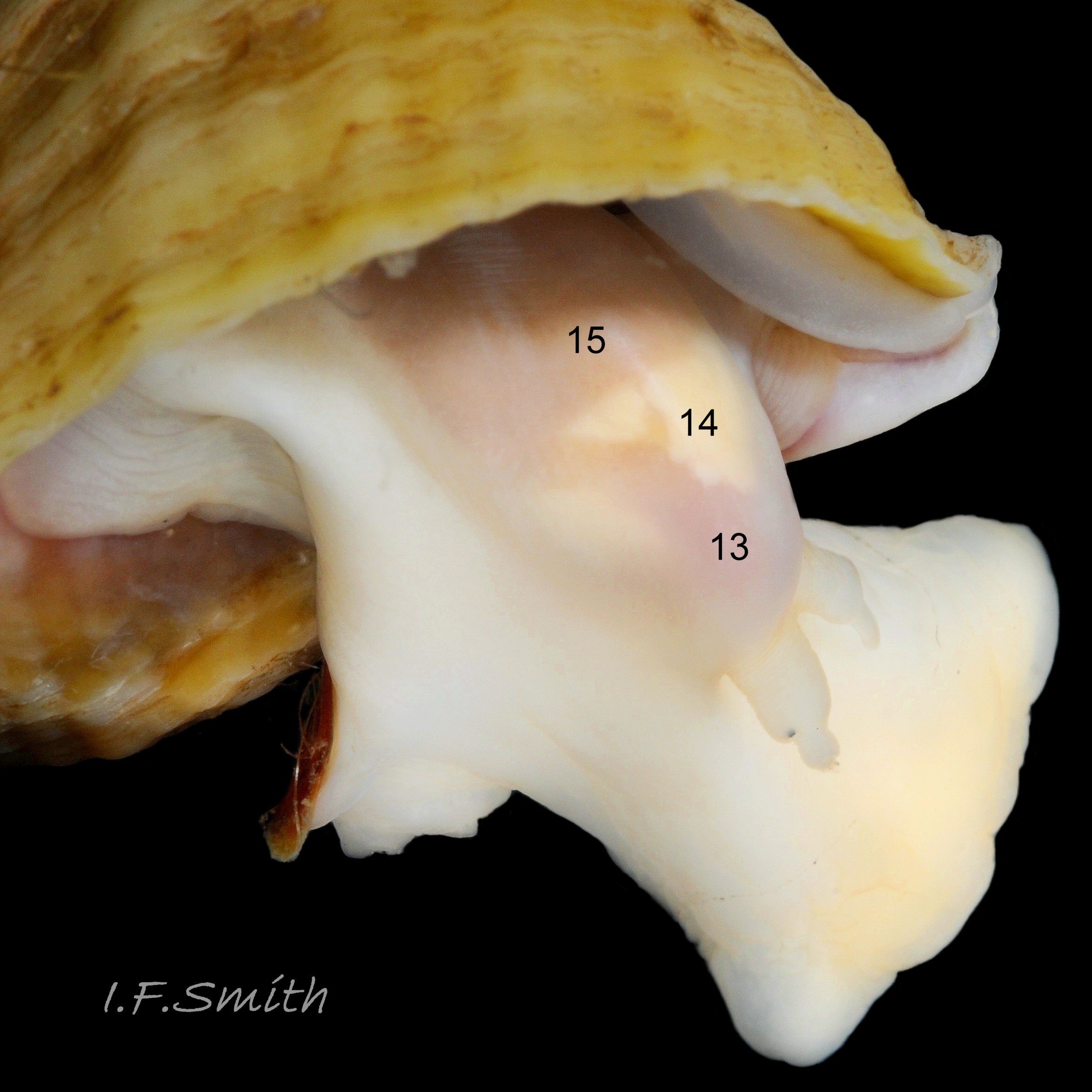
13: buccal mass – pink as rich in myoglobin; contains odontophore and anterior of radula used in rasping prey. (Image 36)
14: acinous salivary gland – compound gland of many small rounded sacs that secrete enzymes for external pre-digestion/ liquefaction of prey. (Image 36)
15: gland of Leiblein – secretes enzymes for internal digestion of ingested liquefied prey. (Image 36)
16: digestive gland – mass of branching tubules and ducts occupying majority of visceral mass (spiral ‘tail’ of body); most visible feature, except in breeding season when much may be covered by ovary or testis. Receives solution of digested food from stomach into tubules where taken up by absorbing digestive cells and passed into blood bathing the tubules. (Images 35a&b, 36, 37, 38)
Reproductive features
17: ovary- gland that produces eggs. (Image 37)
18: albumen gland – translucent whitish gland that secretes albumen for nutrition of developing eggs. (Images 37, 38)
19: sperm ingesting gland – dark brown blind tubules in female where sperm excess to requirements is engulfed and digested by cells; gland is used to establish sex of specimens as penis can occur on females affected by imposex. (Image 37, 38)
20: capsule gland – secretes fibrous wall of capsule containing ova, but does not give it its final shape. (Images 37, 38)
21: testis – gland that produces spermatozoa. (Images 35a&b, 36)
22: prostate – pink gland that secretes fluid which, with spermatozoa and seminal vesicle fluid, forms the semen. (Images 35b, 36)
23: penis – on males, and on females affected by imposex caused by tributyltin pollution. (Image 36)
Key identification features
Nucella lapillus
1. Usual max. height 45mm (to 65mm sublittorally).
2. Shell usually whitish 07 Nucella lapillus , or yellowish 12 Nucella lapillus , some have colour bands 18 Nucella lapillus & 06 Nucella lapillus. No periostracum except on juveniles <3mm high .
3. Short spire and large body whorl with short siphonal canal; white siphon protrudes slightly. (Shells from sheltered shores have longer spires 39 Nucella lapillus, can resemble small Buccinum undatum.)
4. Adults have thick shell with row of teeth inside outer lip of aperture 07 Nucella lapillus. Sub-adults have thin shell and no teeth 06 Nucella lapillus
5.Common on most rocky shores in Britain around mid tide level.
Similar species
Buccinum undatum
39 Nucella lapillus
1. Up to 100mm high (only small ones likely to be confused with N. lapillus).
2. Yellowish brown shell with, often thick, periostracum. Large costae (transverse ribs) curved thus: ) .
3. Large body whorl has short siphonal canal; whitish siphon with black marks (sometimes nearly all-black) protrudes considerably.
4. No teeth inside outer lip of aperture.
5. Common sublittorally all round Britain, sometimes on shore near LWS.
Urosalpinx cinerea
39 Nucella lapillus
1. Up to 40mm high.
2. Yellowish or grey shell with curved costae (transverse ribs).
3. Large body whorl has short siphonal canal.
4. No teeth inside outer lip of aperture.
5. Alien established in Kent and Essex, some expansion of range along S. coast England, but few live records post 2000 A.D. On oyster beds on lower shore and sublittorally.
Habits and ecology
Commonest carnivorous prosobranch on rocky shores in Britain and Ireland. Usually most numerous between MHWN (75% time exposed to air) and MLWN, but up to MHWS if sheltered crevices, and to 20 metres depth in some places. Lives on shores ranging from extremely wave-exposed (Ballantine scale grade1) to extremely sheltered (grade 8), adjusting its low tide behaviour thus: on exposed shores shelters in crevices during storms and active in calm weather, even bright sun; on sheltered shores active at night and in daytime cloudy weather, sheltering in crevices and under rocks from sun. Lower limit of salinity tolerance varies with locality, in Britain usually down to 20 or 25 p.p.t. ; can survive short periods of lower salinity, but not where it is norm. Temperature limits (isotherms of neighbouring oceanic water) up to 19°C in summer, down to 0°C in winter; wider extremes on shore, upper lethal temperature 35°C. Some Swedish populations retreat to sublittoral to avoid winter ice on shore, and in Arctic Russia they retreat under stones on the lower shore. Abundance governed by prey availability; often >10/m² on semi-exposed (grade 4) shores where Semibalanus balanoides (barnacle) abundant, but few on extremely sheltered shores where dense weed-cover inhibits barnacle establishment apart from on occasional steep surfaces 40 Nucella lapillus.
Respiratory water drawn in on left side through inhalent siphon by beat of cilia on robust ctenidium in the mantle cavity. Siphon and pallial channel allow water to be inhaled away from contamination of food, and, with osphradium at inner end of channel, to test water quality and the scents of prey and mates.
Locomotion by ditaxic retrograde waves on centrally divided sole of foot. Slow moving shambling gait. Sedentary as sessile prey usually abundant and unable to flee. Travels about 10cm in a tidal cycle, with days immobile while digests meals. No planktonic stage; dispersal relies on slow crawling; consequent low gene exchange contributes to local differences in appearance and behaviour; chromosome number of races varies.
Eats, in order of usual preference (Crothers, 1985):
Semibalanus balanoides;
Mytilus edulis (mussel, usually under 40mm long) 39a 39a Nucella lapillus ;
Austrominius modestus;
and less than full-grown Perforatus perforatus.
Adults usually ignore small barnacle species such as Chthamalus stellatus & Chthamalus montagui [Some sources contradict Crothers; cause may be geographical variation in genetics or habit, or frequent misidentification by humans of barnacle spp. – examination of tergoscutal flaps required Barnacle Introduction].
When preferred-prey reduced or absent, feed on Patella spp. 40a Nucella lapillus , Littorina spp. or other mollusca. Juveniles up to 8mm high eat tiny Mytilus, Spirorbis (worm) and, in Arctic Russia (Matveeva, 1955), Onoba aculeus (gastropod) in preference to barnacles. In extreme S.W. England, where S. balanoides is rare or absent (settlement and survival vary greatly with weather from year to year, Lewis, 1964, pp.250-251), Mytilus edulis is most frequent prey 39b Nucella lapillus and Patella spp. 40a Nucella lapillus ,Littorina spp. and Gibbula spp. are frequently attacked (Crothers, 1985). Switching to Patella also occurs in areas, such as North Wales and Orkney, where S. balanoides breeds and survives consistently. In Guernsey 31 of 100 vacant Patella shells examined from sheltered shore (Ballantine scale 6-7; near-continuous fucoid cover) had completed Nucella-boreholes (Flint, 2001), suggesting high rate of predation, especially as Patella less than 35mm long can be forced off without boring. On sheltered shores sweep of dense fucoids often hinders settlement of barnacles 40 Nucella lapillus , so reducing favoured prey and causing switch to Mytilus. But no Mytilus observed on this Guernsey shore so Patella the next best thing available. On a very exposed Guernsey shore (Ballantine scale 2; no fucoid cover, barnacles abundant) only 2 of 99 examined vacant Patella shells had completed Nucella-boreholes (Flint, 2001), suggesting less switching to Patella where plentiful barnacles. Oyster spat eaten in Essex. When a prey species exhausted, often delayed transfer, even to ‘preferred’ species, from what locally accustomed to while learns how to handle new prey.
Adults force way between opercular valves of barnacles with proboscis. Paralysing narcotic and digestive enzymes from salivary glands 22 Nucella lapillus injected to liquefy flesh that is ingested through proboscis. Only small shells of M. edulis can be forced apart 39a Nucella lapillus, and small N. lapillus cannot force way into barnacles. In these cases, and on other large prey, N. lapillus bores neat hole through shell 40b Nucella lapillus. by alternating rasping by radula with application by accessory boring organ of enzymes and acid that soften conchiolin and lime in prey’s shell. Angle of radula constantly changed to give smooth round hole; straight rasp-marks show that no rotary action involved 40b Nucella lapillus. Diameter of hole constant; rim not bevelled and no raised boss at base of unfinished hole 40c Nucella lapillus (bevel and boss found in unfinished holes made by Euspira spp. mainly in bivalves) . On limpets, Nucella usually bore into pedal-retractor muscle 40d Nucella lapillus, or central area enclosed by it 40e Nucella lapillus, where radula can reach nutritious viscera; of 73 vacant bored shells in Guernsey only 7% of holes were into viscera-free periphery distal of the muscle, though periphery was 52% of area of shells (Flint, 2001). Boring and feeding take between a day and over a week, depending on temperature and size of prey, followed by resting in sheltered position for day or two after barnacle meal to about five days after a mussel. Boring causes much wear on anterior teeth of radula. Liquid diet results in small amounts of small faecal pellets. Neighbours of assaulted Mytilus can counter-attack by attaching byssus threads to attacker and tightening them to pull it off victim and turn it over so foot is unable to grip substrate to escape 41 Nucella lapillus. This may be targeted, rather than accidental, as non-predatory Littorina littorea on mussel beds rarely so ensnared. N. lapillus seem aware of risk as small mussels without close neighbours preferred to dense beds, and boring usually near posterior adductor muscle away from mussel’s foot and byssus 41a Nucella lapillus. Possible alternative explanation: Littorina littorea more active than N. lapillus so moves and snaps attached byssus threads before numerous and set enough to restrain it. Many free N. lapillus have snapped byssus threads on shell from when moved in time to escape 26 Nucella lapillus.
Consumption of N. lapillus by humans discouraged by acrid smell and taste, but several known predators; list in Crothers (1985) p. 302. Main enemies are Carcina maenas and Necora puber (crabs) and Larus spp. (gulls), and it is one of the hosts parasitized in sequence with Cerastoderma edule and Larus spp. by Parorchis acanthus (trematode worm). Thick shell can sustain considerable abrasion/erosion, but Polydora worms, initially commensal, may eventually cause its decay with multiple borings 01 Nucella lapillus
Variations in shell colour have been attributed to diet (barnacles :white, Mytilus : dark), but environmental selection and genetics seem more likely. Colour of individual’s new growth can change in response to impact of move from shore to aquarium; can confuse laboratory colour: diet experiment results. Populations with large proportions of coloured and banded shells are most frequent in S. Wales and S.W. England, including, but not only, the area where Mytilus consumption predominates. In most populations, old specimens are predominately plain whitish or greyish as outer pattern-bearing layer is eroded away 04 Nucella lapillus.
Variations in shell shape, size, strength and sculpture are related to combinations of environmental pressures and genetic factors. Adults from sheltered shores, when compared with those from neighbouring exposed shores, on average, have longer spires, smaller apertures, and stronger shells 42 Nucella lapillus . These features help resist attack of crabs (common on sheltered shores) by giving space to withdraw into spire, less space for crabs to insert their chelae and harder-to-crack shells (in N. Wales, average force of 580 Newtons to break adult shells on very sheltered shores, 240N on exposed shores). The small aperture also reduces evaporation and the risk of desiccation during low water, and imbrication is more common as there is less erosion to wear it away where there is little wave action. Adults from exposed sites have shorter spires, larger apertures and weaker shells. Here crabs are scarce, and frequent spray reduces the risk of desiccation so the weaker shell and larger aperture are less of a risk, and the larger foot accommodated gives a firmer grip against wave impact. Withdrawal from crabs is less important than the more compact wave-resistant shape formed by the small spire. The exposure related shell differences hold within local areas, but genetic differences affect absolute measurements. In both Pembroke (S. Wales) and the Great Orme (N. Wales) the above relationships hold locally, but on shores of equal exposure (high or low) the Pembroke shells are shorter spired than the Great Orme shells. It seems that the gene pool available in Pembroke lacks the genes needed for very long spires and the pool at the Orme lack those needed for very short spires. This difference is found round Britain; those in the North Sea, north coast of Scotland, Severn Estuary, Liverpool Bay and Kent to Portland being like those on the Orme, and elsewhere shells are like those in Pembroke (map on p.337 in Crothers, 1985).
Breeding occurs all year with a peak in winter or spring (April-May in Yorkshire when water 9°C). Thirty or more congregate in a pool or sheltered crevice for females to be repeatedly inseminated by the males’ long recurved penes, interspersed with periods of laying. An egg capsule passes from the female’s oviduct, along a temporary groove in the foot, to the sole which manoeuvres it into the ventral pedal gland just in front of the centre of the sole. The capsule is held with the plugged opening innermost and the base protruding while the gland squeezes and moulds the capsule into a smooth stalked vase-shape 8-10mm high, 2-4mm wide with a longitudinal seam down two sides. The base of the stalk is pressed into a disc and cemented onto the substrate 43 Nucella lapillus. When foot and gland are lifted off, the conchiolin walls of the capsule harden on contact with sea water. Capsules usually yellow, sometimes mauve or brownish. Female can produce about ten capsules in 24 hours; a breeding congregation may deposit hundreds together. Central mass of capsule made of about 600 agglutinated “nurse” eggs that cease developing at early stage. Ten to thirty unarrested eggs hatch into non-planktonic veliger larvae; small velum with no food-collecting groove; larvae attach to central mass with sucking lips to feed. Temperature controls development time within capsule; four months on S. coast England, five months in Yorkshire, seven months in the White Sea. No free-swimming veliger stage; young emerge from the unplugged apical opening of capsule as crawling snails with globular, smooth, glossy shell (protoconch) of 2½ whorls about 1mm high. Hatching success varies; about 100% of capsules continuously immersed in sea water, about 25% if in cleft or rock pool that receives freshwater run off, 0% to about 60% if at MLWN and dry out at low tide. Hatchlings often shelter in empty barnacle shells. Growth, March-October; rate varies, slower in north than south, in England usually 10-16mm high at 1yr old, 12-26mm at 2yrs, growth usually ceases (but shell thickens) in third year at shell height 20-30mm (more on sheltered shores), but may continue for those castrated by parasites. Some live to over 6yrs old, but mortality heavy before thick shell and mouth bosses develop; about 10% of each cohort survive to 1 year old, 5% to 2yrs, 1.25% to 3yrs.
Reproduction of N. lapillus was greatly reduced in parts of Britain and Europe by imposex where females develop male features including penes that block oviducts and prevent escape of ova. In Plymouth, 5% of females affected in 1970, 67% in 1985; some populations near marinas entirely destroyed. Recognised c.1970, caused by tributyltin in anti-fouling paints on shipping and other marine structures. Tributyltin paint banned on small boats 1987 (UK) and 1991 (EU). Worldwide total ban agreed 2008, ratified by countries accounting for 80% of world shipping by 2012. Recovery of some populations slow; imposex still prevalent where much international shipping. Many other spp. affected world wide; N. lapillus very sensitive to minute amounts so used as indicator sp. for imposex incidence.
Primary function of hypobranchial gland of gastropods generally thought to be trapping in mucus of particles from inhalent water before reach ctenidium, and transporting particles out of mantle cavity. In N. lapillus additional suggested functions of hypobranchial gland have included attraction of mates by scent, and acrid smell/taste to discourage predators. When exposed to light and air, white hypobranchial mucus turns yellow 36 Nucella lapillus and, eventually, purple 44 Nucella lapillus. Exact sequence varies; may include green, reddish, blue and brown 45 Nucella lapillus & 46 Nucella lapillus. Hypobrachial gland mucus on cloth. and sometimes changes little from yellow. Similar mucus from Mediterranean Murex used in Ancient Roman times to dye emperors’ robes purple. Reports of small scale use of N. lapillus to dye cloth in Ireland in 17th Century and, perhaps, Anglo-Saxon England. Several vernacular names derive from ‘purple’. Crushed egg capsules can produce same colours, so probably contain same chemicals; may be cause of capsule colour varying from yellow to mauve or brown.
Distribution and status
White Sea to Gibraltar, not Baltic (low salinity), absent or scarce in Denmark, Germany and Netherlands (soft substrate) . GBIF map www.gbif.org/species/5193449 . Common on hard substrate all around Ireland and Britain, avoiding low salinity of inner estuaries. Apparently absent or scarce in Lincolnshire and Suffolk (soft substrate); many records exist around Ireland and E. Scotland (McKay & Smith, 1979) that have not been entered on NBN. U.K. interactive distribution map NBN https://species.nbnatlas.org/species/NBNSYS0000188539
Some local gaps difficult to explain; some perhaps due to imposex near international ports where tributyltin still present.
Images of Nucella lapillus by other Flickr users:
Nucella lapillus (Arctic Russia)
flic.kr/p/i7Pmg8 (Germany)
flic.kr/p/oBGyCv & flic.kr/p/bqcXVF (Atlantic France)
Nucella lapillus (Galicia, N.W. Spain. Various colours, incl. black)
flic.kr/p/aq43TK & flic.kr/p/d9Rfpw (Massachusetts, USA)
Nucella lapillus (New Brunswick, Canada. Various colour forms)
Acknowledgements
For help and advice about anatomy I would like to thank Dr Gregorio Bigatti, Dr Alfredo Castro-Vazquez, Dr Sami Ibidli, Dr Ivan Nekhaev and Dr Yu I Kantor. Many thanks to Dr Jan Light for the loan of specimens, to Dominic Flint for access to data in his unpublished study and to Paul Challinor and Neil Ward for use of their images. Special thanks for correspondence and patient help are due to Dr Alexandra Richter. Any remaining inaccuracies are attributable to me.
Links and references
The most used sources for this account were Crothers (1985) and Fretter & Graham (1962). Fretter & Graham (1994) contains updated information, but lacks the systematic index of the 1962 edition that enables the finding of N. lapillus details scattered through 800 pages.
Andrews, E.B. & Thorogood, K.E. 2005. An ultrastructural study of the gland of Leiblein of muricid and nassariid neogastropods in relation to function, with a discussion on its homologies to other caenogastropods. J. Mollus. Stud. 71: 269-300. Malacological Society, London. Free pdf at: mollus.oxfordjournals.org/content/71/3/269.full.pdf
Ballantine, W.J. 1961. A biologically-defined exposure scale for comparative description of rocky shores. Field studies 1(3): 1-19. Free pdf at: fsj.field-studies-council.org/media/344345/vol1.3_17.pdf
[Ballantine, pp.16- 18, recognised that his use of indicator species lists was area specific. See Zettler, 2013 for further consideration of this topic.]
Biggam, C.P. 2006. Knowledge of whelk dyes and pigments in Anglo-Saxon England. Anglo-Saxon England 35: 23- 55. Abstract at: journals.cambridge.org/action/displayIssue?iid=1182236
Caldwell, M. Marine pollution and sexual confusion in dog whelks. Free pdf of University College of London poster about imposex, but note that illustrations of “Dog whelks” (N. lapillus) are of Buccinum undatum. www.ucl.ac.uk/~ucbpmbc/downloads/poster.pdf
Carefoot, T. 2016 (date viewed by IFS) A snail’s odyssey; learn about whelks and relatives. [Web-page with detailed information on shell boring by Nucella]
www.asnailsodyssey.com/LEARNABOUT/WHELK/whelFeed.php
Crisp, M., Fine structure of some Prosobranch osphradia. Marine Biology22: 231-242 Abstract at link.springer.com/article/10.1007%2FBF00389177#page-2
Crothers, J.H. 1985. Dog whelks: an introduction to the biology of Nucella lapillus. Field Studies, 6, 291-360. Free pdf at
fsj.field-studies-council.org/media/342851/vol6.2_171_col…
Flint, D. 2001. Unpublished study of Nucella predation on Patella spp. in Guernsey.
Forbes, E. & Hanley S. 1849-53. A history of the British mollusca and their shells. vol. 3 (1853), van Voorst, London. (As Purpura lapillus; Free pdf at archive.org/details/historyofbritish03forb Use slide at base of page to select pp.379-387.)
Fretter, V. and Graham, A. 1962. British prosobranch molluscs. Ray Society, London.
Fretter, V. and Graham, A. 1994. British prosobranch molluscs. Revised and updated edition. Ray Society, London.
Graham, A. 1988. Prosobranch and pyramidellid gastropods. Linnean Society of London.
Hughes, R.N. and Dunkin, S. de B. 1984. Behavioural components of prey selection by dogwhelks, Nucella lapillus (L.), feeding on mussels, Mytilus edulis L., in the laboratory. J. Exp. Mar. Biol. Ecol. 77(1-2) : 45-68. Abstract at www.sciencedirect.com/science/article/pii/0022098184900509
Jeffreys, J.G. 1862-69. British conchology. vol. 4 (1867). Van Voorst, London. (As Purpura lapillus; Free pdf at archive.org/details/britishconcholog04jeffr . Use slide at base of page to select pp.275-289.)
McKay, D.W. & Smith, S.M. 1979 Marine mollusca of east Scotland Royal Scottish Museum, Edinburgh.
Lewis, J.R. 1964. The ecology of rocky shores. London, Hodder & Stoughton.
Mallon, P. & Manga, N. 2007. The use of imposex in Nucella lapillus to assess tributyltin pollution in Carlingford Lough. J.E.H.R. vol.6 issue 2
www.cieh.org/jehr/imposex_nucella_lapillus.html
Matveeva T.A. 1955. Biology of Purpura lapillus (L.) on West Murman. In: Kamshilov M.M., ed. Trudy Murmanskoy Biologicheskoy Stanysii, 2: 48-61 [In Russian].
Medeiros, R., Serpa L. , Brito, C., De Wolf H. , Jordaens, K. , Winnepenninckx, B. & Backeljau T. 1998. Radular myoglobin and protein variation within and among some littorinid species (Mollusca: Gastropoda). Hydrobiologia378: 43-51.
Richter, A., Amor, M.J. & Durfort, M. 2010. The anatomy and ultrastructure of the gland of Leiblein of Bolinus brandis and Coralliophila meyendorfii, two neogastropod species with different ecology and feeding strategies. Poster for Soc. for Environmental Biology, annual meeting, Prague 2010.
Santillo, D., Johnston, P. & Langston, W.J. 2001. Tributyltin (TBT) antifoulants: a tale of ships, snails and imposex. European Environment Agency, environmental issue report 22, part 13.
13. Tributyltin (TBT) antifoulants: a tale of ships, snails and imposex
Sarramégna, R. 1965. Poisonous gastropods of the Conidae family found in New Caledonia. Technical paper 144, S. Pacific Commission, New Caledonia.
www.spc.int/DigitalLibrary/Doc/FAME/Reports/Sarramegna_65…
European Environment Agency. Several articles on imposex and its effects on various species. glossary.eea.europa.eu//terminology/sitesearch?term=imposex
Yonge, C.M. and Thompson, T.E. 1976. Living marine molluscs. Collins, London.
Zettler, M.L. et al. 2013. On the myths of indicator species: issues and further consideration in the use of static concepts for ecological applications Plos One Vol 8, Issue 10 [Ref. is not specific to N. lapillus, see note under Ballantine, above.] Free pdf at
www.ncbi.nlm.nih.gov/pmc/articles/PMC3797757/pdf/pone.007…
Glossary
abapertural = away from aperture.
acinous salivary gland = compound gland of many small rounded sacs that secrete enzymes for external predigestion/ liquefaction of prey.
adapertural = towards aperture.
adapical = towards the apex of the shell.
afferent = carrying towards. (e.g. of vessel carrying blood, see efferent.)
aperture = mouth of gastropod shell; outlet for head and foot.
Ballantine scale = biologically-defined wave exposure scale (see references).
bipectinate = feather-like, with narrow filaments either side of central stalk.
capsule gland = secretes fibrous wall of capsule containing ova.
cephalic = (adj.) of or on the head.
chelae = (singular chela) pincers of crabs and other crustacea.
cilia = (pl.) vibrating linear extensions of membrane used in feeding or locomotion. (“cilium” singular).
ciliated = (adj.) coated with cilia.
columella = solid or hollow axial “little column” around which gastropod shell spirals; hidden inside shell, except on final whorl next to lower part of inner lip of aperture where hollow ones may end in an umbilicus or siphonal canal.
columellar = (adj.) of or near central axis of spiral gastropod.
columellar lip = lower (abapical) part of inner lip of aperture.
columellar muscle = attaches body, including opercular disc, to columella of shell; contraction of muscle withdraws body within shell, and pulls operculum to seal aperture.
commensal = (adj.) obtaining nutrients, shelter, support, or locomotion from a host species, without causing it significant detriment.
conchiolin = horny flexible protein that forms the matrix for the deposition of calcium carbonate to create a mollusc’s shell.
ctenidium = comb-like molluscan gill; usually an axis with a row of filaments either side.
ditaxic = (of locomotion waves on foot) double series of waves, out of phase with each other, one series on each side of central furrow on sole.
direct = (of locomotion waves on foot) waves travel from posterior to anterior.
efferent = carrying away from. (e.g. of vessel carrying blood from ctenidium).
fasciole = (see siphonal fasciole)
gland of Leiblein = secretes enzymes for internal digestion.
height = (of gastropod shells) distance from apex of spire to base of aperture.
hypobranchial gland = thickened, sometimes puckered, tissue on roof of mantle cavity of many gastropods. Emits mucous to trap particles from
inhalent water before it reaches ctenidium. Often other biologically active compounds produced. Gland occurs also in some bivalves and cephalopods (ink sac).
imbricate = shell sculpture of growth-line ornament overlapping like roof tiles.
MHWN = mean high water neap tide level (mean level reached by weakest high tides for a few days every fortnight).
MHWS = mean high water spring tide level (mean level reached by highest tides for a few days every fortnight; Pelvetia zone on rocky coasts).
MLWN = mean low water neap tide level (mean level reached by weakest low tides for a few days every fortnight. i.e. those that fall the least).
mantle = sheet of tissue that secretes the shell and forms a cavity for the gill in most marine molluscs.
mesopodium = middle section of gastropod foot. (see propodium & metapodium).
metapodium = rear section of gastropod foot. (see mesopodium & propodium).
myoglobin – red oxygen-binding protein in muscle tissue; often in buccal-mass muscles of gastropods. Similar to red haemoglobin in vertebrate blood, but green haemocyanin is usual oxygen-carrier in mollusc blood. See www.researchgate.net/publication/251227038_Radular_myoglo…
N = (See Newton).
Newton = (abbreviation N) force exerted by Earth’s gravity on approx. 100g.
odontophore = cartilaginous “tongue” that supports and protracts /retracts the radula.
opercular = (adj.) of the operculum.
opercular disc = part of foot attached to inner face of operculum.
opercular lobe = extension of opercular disc round edge of part of operculum.
operculum = plate of horny conchiolin, rarely calcareous, used to close shell aperture.
osphradium = chemo-receptor organ in molluscs that tests inhalent water flow approaching ctenidium (gill) for “smell” of food, prey, predators, mates and/or water quality.
penes = (plural of penis) male copulatory organs.
periostracum = thin horny layer of chitinous material often coating shells.
plankton = animals and plants that drift in pelagic zone (main body of water).
propodium = front section of gastropod foot. (Cf. mesopodium & metapodium).
prosobranchia = 20th Century term for subclass of Gastropoda that included most marine snails with ctenidia. Now distributed between several subclasses. See note at www.marinespecies.org/aphia.php?p=taxdetails&id=102
rectal gland = (a.k.a. anal gland) Function uncertain, perhaps produces substances that supplement the excretory activity of the kidney.
retrograde = (of locomotion waves on foot) waves travel from anterior to posterior.
sessile = (of organism) fixed in one place, e.g. barnacles.
siphon = extension of mantle to form a channel for inhalent respiratory water current.
siphonal canal = grooved or tubular extension of outer lip of the shell aperture on some snails to support the siphon.
siphonal fasciole = raised rib along columellar side of siphonal canal, bearing curved growth lines formed by successive positions of canal end.
sperm ingesting gland = (in female Nucella lapillus) group of dark brown blind-ended tubules at posterior of capsule gland where excess sperm unrequired by female are trapped, engulfed by cells and digested.
suture = groove or line where whorls of gastropod shell adjoin.
vas deferens = tube carrying sperm to male’s penis.
veliger = shelled larva of marine gastropod or bivalve mollusc which swims by beating cilia of a velum (bilobed flap).